|
|
Územie, ktorého sa dotýka nasledujúci popis, je ohraničené na SZ Bratislavou, odtiaľ smerom na východ pozdĺž úpätia svahov Malých Karpát až k údoliu rieky Váh, ďalej popri Váhu smerom na juh ku Komárnu a odtiaľ poza Dunaj za štátnou hranicou s Maďarskom smerom na Győr a Mosonmagyaróvár, na západe prechádza poza štátnu hranicu s Rakúskom po Bratislavu. Takto vymedzené územie sa rozkladá na Slovensku v Podunajskej rovine a v Maďarsku na Mošonskej rovine. Z geologického hľadiska zahŕňa celú Podunajskú a Mošonskú rovinu (Mosoni síkság), geologicky nazývanú Centrálna depresia Podunajskej nížiny –– Gabčíkovská panva (Obr. III.1).
Toto územie sa nachádza v Dunajskej panve, ktorú na území Slovenska zastupuje Podunajská nížina a v Maďarsku Malá uhorská nížina. Panva je na západnej strane ohraničená Východnými Alpami, Litavským pohorím a na severe Malými Karpatami. Na severe panva vybieha v podobe výbežkov – zálivov – medzi jadrové pohoria Malé Karpaty, Považský Inovec a Tribeč. Na severovýchode sú stredoslovenské neovulkanity a vulkanity pohoria Burda. Na juhovýchode spod panvy vystupujú horniny Zadunajského stredohoria.
Jadrom Dunajskej panvy je centrálna Gabčíkovská panva – depresia. Táto je na slovenskom území ohraničená približne obcami Čunovo – Rusovce – Podunajské Biskupice – tok Čiernej vody – Vlčany – Kolárovo – Kameničná (pri Váhu) – Kližská Nemá – a pokračuje ďalej hlboko cez maďarské územie, Gönyű, poza Mošonský Dunaj – naspäť k Čunovu, Obr. III.2.
Začiatok formovania sa vlastnej Dunajskej panvy siaha do spodného miocénu (pozri Tab. III.1.). Na báze (v centrálnej a východnej časti panvy - spodný báden) sú najmä terestrické a jazerno-riečne sedimenty prechádzajúce do morských sedimentov. V strednom a vrchnom bádene v morskom prostredí sedimentujú íly, uhoľné íly, slojky uhlia, vápnité íly, ílovce a prachovce s vrstvami pieskov a pieskovcov. V sarmatskom období sedimentovali v plytkom brakickom prostredí vápnité íly, prachovce a piesky, v príbrežných častiach zlepence, organodetritické vápence, pieskovce, íly i slojky uhlia.
Usadeniny panonu a pontu sedimentovali v plytkom brakickom jazernom prostredí, v centrálnej časti v hlbšom jazernom prostredí. Na západe sa vyzdvihovali Alpy a na severe Karpaty. Panva sa zapĺňala predovšetkým materiálom z dvíhajúcich sa Karpát, deltovým klastickým materiálom prinášaným pra-riekami. Vrchno-miocénne a pliocénne íly, prachy, a pieskovce dosahujú v centrálnej (gabčíkovskej) depresii hrúbku 2000 – 3500 m (Kováč, 2000). V západnej a severnej časti Gabčíkovskej depresie prevládajú piesky a prachy, vo východnej časti íly a prachy.
Pliocénne usadeniny v Gabčíkovskej depresii sú vekovo zaradené do dáku. Nad nimi je kolárovské súvrstvie zaradené do rumanu (romanu). Sedimenty pochádzajú z paleo riek (Hrona, Nitry, Váhu) a tvoria ich v okrajových častiach piesky, štrky, prechádzajúce smerom do panvy do vápnitých ílov až prachovcov.
Z hľadiska režimu povrchových a podzemných vôd nás zaujímajú hlavne kvartérne sedimenty. Systematickým výskumom kvartéru v Podunajskej nížine sa venovali hlavne na Geologickom ústave Dionýza Štúra v Bratislave. Výsledky výskumov sú zhrnuté v záverečných správach a v publikáciách. Súhrnný prehľad je uvedený napríklad vo „Vysvetlivkách ku geologickej mape JV časti Podunajskej nížiny 1:50000“ (Vaškovský et al., 1982). K poznatkom prispelo i množstvo prác, vykonaných pri inžiniersko-geologickom a hydrogeologickom prieskume pre sústavu vodných diel Gabčíkovo-Nagymaros. V ostatnej dobe komplexný pohľad na geologické pomery z rôznych hľadísk a najnovšie poznatky podáva Záverečná správa „Podunajsko – DANREG“ vypracovaná pod záštitou Ministerstva Životného prostredia SR v roku 1996 (Tkáčová, Kováčik et al.), stručne citovaná ako DANREG a Mapa a Vysvetlivky k neotektonickej mape Slovenska, Maglay, 1999. Z principiálneho hľadiska tvorby sedimentov je veľmi poučná publikácia Michalíka et al. 1999, Geológia stratigrafických sekvencií.
III.1.1 Geologická definícia územia (DANREG, 1996, 2000)
Pokiaľ riešime praktické úlohy prúdenia podzemných vôd budeme sa z geologického hľadiska zaoberať predovšetkým kvartérnymi sedimentmi, z ktorých najdôležitejšie sú náplavy riek v riečnych a jazerných sedimentačných podmienkach, a to predovšetkým Dunaja a prítokov z Karpát (Váh, Nitra, Hron).
Tab. III.1 Schéma geologického členenia treťohorných a štvrtohorných útvarov
(spracované podľa Kováč, 2000; Michalík, 1999; Hovorka, Michalík, 2001)
|
|
Územie monitorovania prírodného prostredia patrí do Centrálnej depresie Podunajskej panvy. Voči Malým Karpatom je Centrálna depresia Podunajskej panvy výrazne tektonicky ohraničená okrajovými malokarpatskými dislokáciami. Zo SV strany ju voči Trnavskej a Nitrianskej pahorkatine ohraničuje systém zlomov SZ-JV smeru. Tieto zlomy usmerňujú zväčšovanie hrúbok kvartérnych sedimentov smerom do centra depresie a ohraničujú depresiu voči vyšším kryhám východnej časti Podunajskej nížiny. Od SZ okraja (od Malých Karpát) báza kvartéru Centrálnej depresie Podunajskej nížiny prudko klesá po obce Bodíky, Šuľany, Horný Bar do hĺbky 450 – 500 m (najväčšie zistené hĺbky na vrtoch sú pri Gabčíkove 450, 459 a 468 m). Zdá sa, že na maďarskej strane oproti Bodíkom a Vojke dosahuje hĺbka kvartéru až 600 m, hoci priamo vrtom bola zistená hĺbka najviac 414 m. Podložie potom stúpa smerom k východnému okraju depresie. V kvartérnej výplni depresie sú tri výrazné súvrstvia.
Vrchné súvrstvie, len niekoľko metrov hrubé, prevažne jemnozrnné sedimenty holocénu.
Stredné súvrstvie so stredne až hrubopiesčitými štrkmi s ojedinelými medzivrstvami jemnozrnnejších sedimentov.
Spodné súvrstvie tvoria prevažne drobnozrnné piesčité štrky s častými medzivrstvami ílovitých alebo siltovitých pieskov až ílov.
Podložie kvartéru tvoria sedimenty rumanu, dáku a pontu, spoločne nazývané ako pont – pliocén. Počiatkom pontu sa polobrakické more naďalej vysladzovalo a Podunajská panva sa zmenila na okrajový záliv, v ktorom počas pliocénu sedimentácia prebiehala už v sladkovodných jazerno – paludálnych podmienkach a od rumanu (najvyšší pliocén) už v riečno-jazerných podmienkach. Tieto prechádzajú postupne do spodného súvrstvia kvartéru.
V centre Gabčíkovskej depresie sú prevládajúcim typom sedimentov pont-pliocénu sivé až zelenosivé, slabo vápnité, sľudnaté íly a prachy s premenlivou piesčitou prímesou. Striedajú sa s polohami jemne až strednozrnných, slabo vápnitých pieskov, lokálne stmelených do pieskovcov. Okrajové fácie obsahujú hrubozrnnejší materiál pochádzajúci z jadrových pohorí. Tieto sa, z hľadiska prúdenia podzemnej vody v kvartérnych sedimentoch, považujú za nepriepustné podložie kvartérnych riečnych a jazerných sedimentov.
Pre oblasť Podunajskej nížiny je počas kvartéru charakteristické neustále poklesávanie územia. Zjednodušená geologická štruktúra je nasledujúca. Pod pokryvnou ornicou, piesčitými hlinami, pieskami a zahlinenými štrkami o hrúbke do niekoľko metrov sa nachádza komplex štrkopiesčitých sedimentov. Tento je možné rozdeliť na základe elektrického odporu na hrubozrnné štrkopiesky s vyššou rezistivitou a štrkopiesky s väčším obsahom piesčitej zložky s nižšou rezistivitou, ale stále významnou priepustnosťou. Pod nimi sú horniny s jemnejšou zrnitosťou.
Mapa hrúbky kvartérnych sedimentov v programe DANREG bola spracovaná na základe výsledkov interpretácie VES pre celý región Podunajska (Obr. III.3). Pre Podunajskú nížinu je charakteristické neustále poklesávanie bázy a terénu počas kvartéru a to v centrálnej časti na maďarskom území až 600 m. Najmenšie hrúbky štrkopiesčitých náplavov, do 10 m, sú interpretované v oblasti Bratislava – Rača – Pezinok, na styku Podunajskej nížiny s Malými Karpatami, kde je územie výrazne tektonicky ohraničené malokarpatskými dislokáciami. Zo SV strany je územie ohraničené systémom zlomov smeru SZ – JV voči Trnavskej a Nitrianskej pahorkatine.
Kvartérno-geologický vývoj územia bol podmienený formovaním mohutného toku Dunaja a vo východnej časti jeho prítokmi Váhu, Nitry, Žitavy, Hrona. Kvartérnu sedimentačnú výplň a pokryvy územia tvoria hlavne fluviálne sedimenty riek, prechodné fluvio-limnické súvrstvia, proluviálne a deluviálne sedimenty úpätí pohorí, pokryvy spraše, sprašových hlín, naviate piesky a pôdne horizonty. Kvartérne sedimenty sa v území začleňujú do obdobia spodného, stredného a vrchného pleistocénu a do obdobia holocénu.
Podunajská nížina je súčasťou Podunajskej (komárňanskej) panvy. Dominujúce postavenie v kvartéri majú fluviálne a fluvio-limnické sedimenty, tvoriace sedimentačnú výplň centrálnej depresie Podunajskej panvy. Vyvinuté sú superpozične. Okrem fluviálnych sedimentov sú vyvinuté eolické spraše a piesky, organogénne sedimenty, slatiny a na okrajoch deluviálne, eluviálne a deluviálno-fluviálne sedimenty.
Za najstaršie vrstvy pleistocénu v Podunajskej nížine sú považované limnické a fluviálno-limnické sedimenty, ktoré tvoria výplň najviac poklesnutej centrálnej (Gabčíkovskej) depresie. V Podunajskej panve ležia na rozličných členoch vrchnej stavby neogénu. Kryjú sa s rozsahom kolárovských (gabčíkovských) vrstiev (pieskov, siltov). Tento komplex sedimentov je charakteristický cyklickým striedaním pestrých piesčito-štrkovitých sedimentov s častými ílovitými a hlinitými polohami.
Pre podunajskú nížinu v strednom pleistocéne (mindel) je už charakteristická rozsiahla, prevažne riečna sedimentácia Dunaja a jeho prítokov Váhu, Nitry, Žitavy, Hrona, ktorá leží na podložnom fluviolimnickom spodnom pleistocéne. Panvový vývoj pokračoval synsedimentárnym poklesávaním. Oblasť Váhu bola stabilnejšia. Oblasť Nitry, Žitavy, Hrona a Ipľa boli mierne vyzdvihnuté. V strednom pleistocéne došlo už k definitívnej úprave smerov prúdenia hlavných tokov, v tejto oblasti hlavne Dunaja a Váhu. V tomto období Dunaj a Váh vytvára obrovské ploché vejáre náplavových kužeľov, zakryté pokračujúcou fluviálnou sedimentáciou vrchného pleistocénu a holocénu.
Medzi spodným a stredným pleistocénom je vo východnej časti gabčíkovskej depresie vyvinutá hlinito-ílovitá poloha (koniec plošnej limnickej sedimentácie). V okrajových častiach Podunajskej nížiny fluviálne piesčité štrky stredného pleistocénu ležia na vrstvách kolárovskej formácie, na palárikovských vrstvách a na vrstvách dáku.
Tento komplex stredného pleistocénu sa často označuje ako „dunajská štrková séria“. Jeho hrúbka je pod Bratislavou 10 – 20 m, pri Komárne 8 – 12 m, Sládkovičove 18 – 24 m a v strede depresie v okolí Gabčíkova dosahuje hrúbku cca 160 m. Ide prevažne o hrubšie štrky, piesčité štrky a piesky bez jemnozrnných frakcií, čo je odrazom prevládania korytových fácií sedimentov nad nivnými. Štrkový materiál má charakteristické hrdzavohnedé, hnedožlté a sivé povlaky. V hĺbke 20 – 26 m a 60 – 80 m sa nachádzajú piesčité, prachovité, ílovité hliny a íly pravdepodobne ris-wurmského a mindel-riského interglaciálu.
Na území Žitného ostrova je vrchný pleistocén tvorený fluviálnym štrkopiesčitým súvrstvím (dnová akumulácia tokov) o hrúbke do 15 m. Okrem toho sú tu fluviálno eolické a eolické sedimenty a organogénne sedimenty. Proluviálne sedimenty (ploché náplavové kužele malokarpatských tokov - wurm) tvoria úzky lem na úpätí Malých Karpát a medzi Bratislavou a Modrou.
Na území Žitného ostrova ide hlavne o fluviálne vrstvy prechodného obdobia medzi pleistocénom a holocénom. Vtedy sa vytvorili staršie agradačné valy, hlavne vyvýšené jadro Žitného ostrova tiahnúce sa od Podunajských Biskupíc až ku Komárnu, ktoré sa ďalej ponára pod sedimenty holocénu. V hornej časti šírka holocénu dosahuje 15 km a v dolnej 4 – 6 km, prípadne vystupuje ostrovčekovite. Vyvýšené jadro sa skladá z piesčitých štrkov stredného fluviálneho súvrstvia a z dnovej akumulácie Dunaja. Agradačné valy sa skladajú z jemno až strednozrnných pieskov a dosahujú prevýšenie nad terénom do 3 – 4 m.
Holocénne sedimenty tvoria povodňový nivný kryt riečnych nív, v tomto území Dunaja a Váhu. Tvorené sú striedaním piesčitých až ílovitých hlín, občas s prímesou štrku, premiestnené spraše, sprašové hliny, recentné a fosilné pôdy. V depresiách a v opustených mŕtvych ramenách sa vytvorili a pretrvali slatiny (Jurský Šúr, Modranský Šúr, slatinisko pri Pustých Uľanoch, pri obci Hájske, a iné drobné lokality).
Centrálna depresia Podunajskej panvy má poklesovo – priehybovú stavbu s poklesmi pozdĺž zlomov. Depresia má misovitú brachysynklinálnu štruktúru, vytvorenú bez ohľadu na pred-panónske podložie (Priechodská, Vass, 1986). Zlomy existujú hlavne v starších členoch výplne a sú hlavne voči bádenu synsedimentárne. Tieto zlomy v panóne a v mladších sedimentoch vyznievajú, prípadne sa výšky skokov redukujú na niekoľko metrov. Staršími sedimentmi prechádzajú zlomy SV smeru a vytvárajú tak výraznú blokovú stavbu, poprvé opísanú Adamom a Dlabačom (1961). Zlomové systémy, staršie i mladšie, sa iba v malej miere podieľajú na formovaní Centrálnej depresie Podunajskej panvy. Táto vznikla v panóne a vyvíjala sa až do konca pliocénu. Išlo o pokles hlavne prehýbaním, v malej miere s poklesmi po zlomoch, a to hlavne po okrajoch depresie (Franko et al., 1984).
Hydrogeologické celky v tejto oblasti, hlavne vzhľadom na výskyt termálnej podzemnej vody (geotermálnu energiu), vyčlenil Franko et al. (1984, 1989). Najvrchnejší celok predstavujú štrky, piesčité štrky a piesky rumanu a kvartéru. Miestami sú prítomné tenké nesúvislé vrstvy ílov, hlín, šošovky slatín, a občas aj výplne starých mŕtvych ramien. Vrtmi zistená hrúbka týchto sedimentov je 450, 459, 462 m (pri Gabčíkove), 468 m pri Boheľove, prípadne aj viac, Obr. III.2, Obr. III.4.
Sedimenty rumanu (najvyšší pliocén) vznikli ako jazerné a riečno-jazerné sedimenty, zložené z materiálu, ktoré priniesli staré rieky z dvíhajúcich sa pohorí. Ich typickým príkladom je na sledovanom území Kolárovská formácia (začínajúca v najmladšom pliocéne – rumane?) s materiálom prineseným starou Nitrou a starým Váhom, a ďalej, gabčíkovské piesky (gabčíkovskej depresie) s materiálom prineseným do jazerných sedimentačných podmienok starým Dunajom. Nad nimi pokračovala sedimentácia v kvartéri naďalej jazernými a jazerno-riečnymi sedimentmi najstaršieho pleistocénu a približne od mindelu do holocénu sedimentovali riečne sedimenty tvorené už dnešnou riekou Dunaj (Vaškovská, 1986) a vo východnej časti územia riekami Váh a Nitra. Na území Podunajskej nížiny existovali súčasne od rumanu dve sedimentačne rozdielne podmienky, a to západná časť pod Bratislavou až po približne líniu Veľký Meder (Čalovo) – Šaľa a východná časť od tejto línie po Podunajskú pahorkatinu (Obr. III.4). V západnej časti prevládali sedimentačné podmienky s väčším sklonom dna a terénu a sedimentovali prevažne splaveniny (sedimenty sunuté po dne) Dunaja, vo východnej časti územia bol sklon územia podstatne menší a sedimentovali prevažne sedimenty riek Nitry a Váhu. V západnej časti prevládajú do mindelu preto sedimentačné podmienky viac prietočných jazier a riečno-jazerné a neskoršie riečne, a vo východnej časti prevládajú sedimenty jazerné, sedimenty plytkých jazier s občasnými výskytmi riečnych štrkov. Takéto rozdielne podmienky prevládajú prakticky až do holocénu. Hranica medzi týmito sedimentačnými podmienkami sa menila. Vo východnej časti Dunaj občas oderodoval časť sedimentov a zanechal tam polohy svojich sedimentov a preto i vlastnosti sedimentov sa z miesta na miesto a v závislosti na hĺbke môžu meniť vo veľkom rozsahu. Sedimenty, ktoré sa usadzovali v západnej časti, majú preto podstatne iné vlastnosti parametrov vstupujúcich do hydraulických výpočtov a modelov prúdenia podzemných vôd, ako sedimenty východnej časti (Kolárovskej formácie).
V podloží tohoto celku (rumanu a kvartéru) leží komplex menej priepustných kolektorov a izolátorov – dák – o hrúbke 55 až 1174 m (Franko et al., 1989). Tento komplex považujeme za hydrogeologické podložie štrkopiesčitých sedimentov, ktoré nás v Podunajskej nížine zaujímajú z hľadiska modelovania a riešenia úloh spojených s podzemnou vodou. Všeobecne dák zahŕňa striedanie pieskov a ílov, v jeho najvrchnejšej časti sú hlavne piesčité íly a íly. Pri Senci a Chorvátskom Grobe sa nachádzajú štrky. V strednej časti depresie, ktorá nás najviac zaujíma, štrkové polohy v dáku sa nenachádzajú, striedajú sa tu piesky s ílmi, pričom v niektorých vrtoch vo vrchnej časti dáku prevládajú íly (Franko et al., 1984). Podložie v okolí Bratislavy tvoria granitoidné horniny.
Neotektonickou stavbou Žitného ostrova a priľahlého územia Podunajskej nížiny, hlavne z hľadiska prieskumu podzemných vôd kvartérnych sedimentov, sa zaoberal Pospišil et al., (1978) a Pospišil, Melioris (1981). Na základe prvej významnejšej polohy ílu vo vrtoch, ktorú považovali za bázu komplexu štrkopieskov, vymedzili sieť zlomov, ktoré boli aktívne v kvartéri. Línie medzi blokmi s nerovnakou hrúbkou štrko-piesčitého komplexu pokladajú za línie disjunktívneho charakteru. Počet takto vyčlenených zlomov v práci Pospišila et al., (1978) je okolo 50. Keďže ide o porovnávanie hĺbok podložia štrkov, tieto línie by mali naznačovať existenciu zlomov (alebo zosunov, prípadne dnovú riečnu eróziu) v sedimentoch ich podložia. Sami autori zdôrazňujú, že to neznamená, že pod touto hĺbkou sa už piesok a štrk nevyskytuje. Takýto prístup nevylučuje možnosť vymedzenia rôznych, vzájomne nekorešpondujúcich hĺbok vo vrtoch hlavne preto, že íly tvoria nesúvislé polohy. Takto zistená „báza štrkov“ pravdepodobne predstavuje hranicu medzi riečno-jazernými (sedimentácia riečneho štrku a piesku v jazerných podmienkach) a riečnymi sedimentmi Dunaja (fluviálne štrky rieky Dunaj). Okrem toho, medzi jazernými a riečnymi sedimentmi bola často zistená erózia a preto takáto zistená hranica pravdepodobne len málokedy poukazuje na neotektonické procesy. Významnou pomôckou pri definovaní hranice medzi sedimentáciou v jazerných a riečnych podmienkach je skutočnosť, že v riečnych podmienkach existovali oxidačné podmienky a preto sú štrky potiahnuté povlakom limonitov okrovej a šedej farby. V jazerných podmienkach, v ktorých existovali viac redukčné podmienky, sú sedimenty šedozelenkavej farby s väčším obsahom jemnozrnnej frakcie (silt, íl), čo sa prejavuje aj v ich menšej priepustnosti. Takto interpretovanú neotektonickú stavbu Žitného ostrova nie je možné prijať pre výpočty prúdenia podzemnej vody, ani pre prácu s návrhom diskretizácie modelu prúdenia podzemných vôd už aj preto, že nerozlišuje štrko-piesčité sedimenty fluviálneho prostredia a sedimenty fluviálneho pôvodu usadené v limnických podmienkach.
Výsledky najnovších neotektonických štúdií predstavuje Mapa a Vysvetlivky k neotektonickej mape Slovenska, Maglay, 1999. Túto mapu a pre maďarské územie mapu z DANREGu (2000) doplnenú o údaje z vrtov, berieme za základ ďalších prác, Obr. III. 2, Obr. III.4.
Neotektonický panvový pokles (Gabčíkovská panva) je na mape zvlášť vyznačený a oddeľuje panvové neotektonické kryhy a štruktúry od extrapanvových. Panvové zlomy (zlomy vo vnútri „živej“ panvy) sa vytvárajú ako zlomy panvového typu, pri ohýbaní panvy. Ohýbanie panvy zachovalo prvotný charakter vnútorných zlomov v panve. Zlomy sú v hrubozrnnejších a nespevnených sedimentoch menej výrazné a majú malý sklon. Štruktúry sú zväčša extenzívne a diferencovane poklesové. Vo vnútri vlastnej Gabčíkovskej panvy sa geologicko-geofyzikálnym prieskumom pre obdobie posledných 0,7 mil. rokov nezistila žiadna aktivita a diferenciácia pozdĺž zlomov.
Na okrajoch vlastnej Gabčíkovskej panvy sa nachádzajú prechodné štruktúry s doloženými mladšími pleistocénnymi a holocénnymi až recentnými aktívnymi neotektonickými zlomami a štruktúrami. Neotektonické mimopanvové zlomy tvoria systém relatívne zdvihových a poklesových štruktúr.
V panóne more pokrývalo územie smerom na SZ až po Viedenskú bránu nad Viedenskou panvou (Fink, 1966). Dunaj v Alpách, už ako väčšia rieka s veľa prítokmi, vznikol vo vrchnom pliocéne – rumane a postupne sledoval ustupujúce more. To súvisí s tektonickými pohybmi, hlavne dvíhaním sa Álp a Karpát. Posledná orogénna fáza na prechode pliocén – pleistocén je zvlášť dôležitá. Za ustupujúcim morom sa začína koncom vrchného pliocénu v budúcej Podunajskej nížine formovať riečna sieť. Vznikajú sladkovodné prietočné jazerá a v nich sedimentujú nánosy prinášané Dunajom a bočnými prítokmi, hlavne od dvíhajúcich sa Karpát, pra-Váh, pra-Nitra. Pôvodne pra-Dunaj vtekal do Podunajskej nížiny Korutánskou bránou, neskoršie preráža pohorie Malých Karpát pri Devíne, v mieste, kde prebiehajú priečne zlomy, medzi ktorými sa vytvorila malá zníženina zvaná Devínska brána. Toto sa udialo vo vrchnom pliocéne – rumane.
Koncom vrchného pliocénu (koncom rumanu) Dunaj nachádza cestu do Veľkej maďarskej nížiny medzi Nagymarosom a Vyšehradom. V strednej a dolnej časti Podunajskej nížiny vznikajú riečno – jazerné podmienky sedimentácie s litorálnou sedimentáciou hrubších sedimentov a sublitorálnou sedimentáciou jemných pieskov až bahna, čo bolo závislé na rýchlosti prúdenia vody v jazerách. Spočiatku sa to deje ako výsledok ústupu panonského mora a neskoršie v súčinnosti s dvíhaním sa Maďarského stredohoria. Dôkazom môžu byť terasy nad Dunajom pri Ostrihome. Relatívne poklesávanie Podunajskej nížiny trvá aj v kvartéri, avšak od mindelu bez riečno-jazerných podmienok (len fluviálne - riečne podmienky), pretože Dunaj stačil vypĺňať relatívne poklesávajúce územie štrko-piesčitými nánosmi, ba dokonca vytvoril mohutný náplavový kužeľ, prevyšujúci okolitý terén. Napriek tomu sa v týchto riečnych náplavoch zachovali hydromorfné pôdy a v niektorých prípadoch boli v štrkoch zistené rašeliny.
Moment prechodu od jazerno – riečnych podmienok k riečnym podmienkam sedimentácie v mindeli (Tab. III.1) je dôležitý z hľadiska priepustnosti, vrstevnej heterogenity i filtračnej anizotropie zvodneného štrko-pieskového prostredia. Zároveň je dôležitý i z hydrogeochemického hľadiska, pretože vzhľadom na zrnitostné zloženie a obsah organických látok sa pri sedimentácii v riečno-jazerných podmienkach mohli vytvoriť a udržať redukčné podmienky.
Zaujímavé je z takéhoto hľadiska rozloženie geologických profilov hlbokých vrtov, ktoré sú uvedené v prácach Franko et al. (1984) a Janáček (1967). Vidieť, že práve sedimenty dáku a rumanu odpovedajú sedimentom pra-Dunaja, sedimentujúcim do jazerného prostredia. Tieto sa v prácach Janáčka nazývajú gabčíkovské piesky. Ich sedimentáciu v jazernom prostredí potvrdzuje zrnitostné zloženie. Riečne dunajské štrky sú veľmi hrubé, o priemere až 10 – 17 cm. Sedimenty gabčíkovských pieskov sú tvorené drobnými štrčíkmi až strednozrnnými pieskami. Okrem toho dunajské riečne štrky sú obyčajne zafarbené limonitom, dokazujúcim oxidačné vodné prostredie. Gabčíkovské piesky a drobný štrčík sú šedé až zelenošedé, bez povlakov limonitu, čo hovorí o redukčnom prostredí v jazernej kľudnej sedimentácii. Medzi jazernou sedimentáciou gabčíkovských pieskov a dunajských štrkov je zrejmá diskordancia (Janáček, 1967) a je zrejmý intenzívny odnos (riečna erózia) po ústupe jazier. V Podunajskej nížine teda existujú riečne aluviálne sedimenty Dunaja a jeho prítokov, hlavne Váhu a Nitry, a riečne sedimenty Dunaja a jeho prítokov sedimentujúce do jazerného prostredia. Okrem toho existujú sedimenty litorálneho a sublitorálneho pásma. Na to všetko vplýva sklon územia a zmeny relatívneho poklesávania stredu panvy a tiež petrografické zloženie hornín z ktorých rieky prinášali materiál. Všetky tieto vplyvy sa v konečnom dôsledku prejavia v priepustnosti a v zrnitostnom zložení šrtkopiesčitého priestorového komplexu zvodneného prostredia v závislosti na polohe a hĺbke.
Súbežne s gabčíkovskými pieskami začala na východ od nich vznikať v rumane Kolárovská formácia, sedimentujúca v kľudnejšom plytkom jazernom prostredí so znosom materiálu z oblasti Karpát.
V záujmovom území na západnej strane existuje v rumane jazerná a následne jazerno riečna sedimentácia gabčíkovských pieskov a štrčíkov, sedimentovaná hlavne pra-Dunajom a karpatskými paleotokmi. Súčastne, na východnej strane, existuje jazerná sedimentácia kolárovských vrstiev, sedimentovaná hlavne karpatskými pra-tokmi (Váh, Hron). Toto sa deje až po obdobie mindelu.
Nástup mindelu bol sprevádzaný zdvihovou a poklesovou tektonickou tendenciou. V západnej oblasti začína prevládať typická dunajská riečna sedimentácia mohutného náplavového kužeľa. Na východnej strane sa Dunaj posúva na sever a východ a začína tiecť do Veľkej maďarskej nížiny územím, ako ho poznáme dnes. Približne od línie Veľký Meder (Čalovo) – Čičov sa zachováva podstatne menší sklon rieky a približne ostávajú zachované predchádzajúce sedimentačné podmienky. Pokračuje sedimentácia kolárovskej formácie riečnymi sedimentmi, hlavne Váhu, miestami striedané so sedimentami Dunaja, a to v podmienkach prúdenia s malým sklonom, často v plytko jazerných a jazerno-riečnych podmienkach. Hranica medzi dunajskými riečnymi sedimentmi a sedimentmi kolárovskej formácie je pomerne ostrá. Hydraulické vlastnosti zvodneného prostredia dunajských riečnych sedimentov a sedimentov kolárovskej formácie sú výrazne rozdielne.
Na základe hlbokých geotermálnych vrtov (Franko et al., 1984) a hlbokých vrtov (Janáček, 1967) a ďalších vrtov, ktoré dosiahli podložie (Obr. III.2), prípadne hranicu medzi jazernými a riečnymi sedimentmi, sme vypracovali (Mucha et al., 1991) mapu izolínií (v kótach) bázy riečno – jazerných sedimentov a izolínií bázy riečnych sedimentov (Obr. III.4.). Na obraze je zároveň vykreslená hranica medzi dunajskými riečnymi až riečno-jazernými sedimentami a jazernými sedimentami Kolárovskej formácie. Táto hranica, prechádzajúca Veľkým Mederom, je približná, pretože oba typy sedimentácie sa cez ňu vzájomne striedali a prelínali. Mapa izolínií bázy riečno-jazerných a riečnych sedimentov zároveň určuje spodnú hranicu zvodneného prostredia a geometricky definuje ohraničenie zvodneného prostredia. Pre ďalšie výpočty a modelovanie ich považujeme za izolínie nepriepustného podložia.
Z hľadiska koncepčného modelu prúdenia podzemnej vody Žitného ostrova hrá základnú úlohu geometria podložia (pont-pliocén), v strednej časti územia tzv. uholná séria (alebo šedá séria pontu, miocén). Táto séria sa skladá hlavne zo svetlošedých a svetlonazelenalých vápnitých ílov, piesčitých ílov a uhoľných lignitov.
Na uhoľnej sérii sedimentovali v oblasti hornej časti Žitného ostrova, západne od Medveďova - Veľkého Medera, tzv. gabčíkovské piesky a štrky dunajského pôvodu sedimentované v jazerných podmienkach a to od pliocénu (napr. označené ako levant) po stredný pleistocén - mindel. Od mindelu sedimentujú na Gabčíkovských pieskoch a štrkoch pra-dunajského pôvodu už fluviálne dunajské štrky. Smerom k Bratislave dunajské sedimenty ležia na rozličných členoch vrchného neogénu a pri Bratislave aj na žulovom podloží.
Na uhoľnej sérii v oblasti východne od Medveďova – Veľkého Medera sedimentuje piesčito-štrkovitá séria Kolárovskej formácie (silty, piesky, štrčík s ilovitými a hlinitými polohami) sedimentujúcej v jazerných a riečno-jazerných podmienkach. Od stredného pleistocénu v tejto oblasti vytvára hlavne Váh plochý náplavový kužeľ s veľmi malým pozdĺžnym spádom (na rozdiel od náplavového kužeľa Dunaja v západnej časti územia). Od stredného pleistocénu sa často sedimenty Váhu a Dunaja vzájomne na svojom styku prekrývajú.
Podobná stavba je ďalej na východ aj na prítokoch starých riek Nitry, Žitavy, Hrona a podobne aj na juhozápadnej strane Dunaja hlavne juhozápadným smerom od Mošonského ramena Dunaja (Lajtha, Rába, Rábca).
Základným rozdielom medzi sedimentmi Dunaja a jeho prítokov je okrem skladby sedimentov hlavne priepustnosť. Priepustnosť štrkov Dunaja je 10 – 1000 násobne väčšia ako priepustnosť sedimentov prítokov.
Podľa E. Mazúra a M. Lukniša (1986) je záujmové územie súčasťou geomorfologického celku Podunajská rovina, oblasti Podunajská nížina, subprovincie Malá dunajská kotlina, provincie Západopanónska panva. Je mladou štruktúrnou poriečnou rovinou, ktorej vývoj prebieha i v súčasnosti.
Z geologicko-morfologického hľadiska je dôležité, že v kvartéri od mindelu vývoj prebiehal už bez riečno-jazerných podmienok a Dunaj meandroval prakticky od Malého Dunaja po Mošonské rameno Dunaja, rozvetvoval sa, prinášanými štrkopiesčitými splaveninami stačil vypĺňať relatívne poklesávajúce územie, ba dokonca vytvoril mohutný náplavový kužeľ, prevyšujúci okolitý terén po ktorého vrchole aj najviac tiekol Obr. III. 5.
Tak vznikli geomorfologicky výnimočné podmienky. Dunaj tečie po hrebeni svojho vyvýšeného náplavového kužeľa, a nedrénuje svoje riečne sedimenty, ako to rieky obyčajne robia, ale naopak, napája ich svojou vodou. Keď prídu povodne rozlieva sa, zatápa široké územie, a keby sa protipovodňové hrádze roztrhli, záplavy by sa dostali až k Malému Dunaju a Mošonskému ramenu Dunaja. Príkladom môžu byť povodne v roku 1954 a 1965, Obr. III.6a, Obr. III.6b.
Koncom pleistocénu a začiatkom holocénu sa vytvorili staršie agradačné valy, hlavne vyvýšené jadro Žitného ostrova, tiahnúce sa od Podunajských Biskupíc až ku Komárnu. Toto tvorí na mape povrchovú rozvodnicu, ktorá však v skutočnosti nefunguje pre povrchovú a podzemnú vodu ako rozvodnica. Počas zrážok sa tu nevytvára povrchový odtok a zrážková ani podzemná voda za žiadnych vodných stavov na Dunaji neprúdi z územia medzi Dunajom a vyvýšeným jadrom Žitného ostrova do Dunaja, ale smerom do Malého Dunaja. Skutočnou rozvodnicou je Dunaj s ramennou sústavou a dunajská voda napĺňa nielen podzemné vody Žitného ostrova ale aj Malý Dunaj a Mošonský Dunaj. Navrhovať územie Žitného ostrova a Malého Dunaja do povodia Váhu je jednoznačne nesprávne. Nie Dunaj a Malý Dunaj tečú do Váhu ale Váh tečie do ramena Dunaja a z neho do Dunaja. Malý Dunaj bol v minulosti jedným z hlavných korýt-ramien Dunaja a aj plavebnou dráhou. Súčasný Dunaj (a starý Dunaj od Čunova po Sap) je umelé vodné teleso (Directive, 2000).
Pretože územie bolo v celom období štvrtohôr relatívne poklesávajúce, a celý čas bol vysoký prínos štrkov Dunajom, hrúbka jazerno-riečnych sedimentov a náplavového kužeľa v najhlbšom mieste dosahuje okolo 500 m. Pri každom vodnom stave na Dunaji a zvlášť pri vysokom vodnom stave hladiny vody, dunajská voda vteká do mohutného náplavového kužeľa, zvyšuje sa hladina podzemnej vody, ktorá pri dlhodobom zvýšení hladiny môže vystúpiť nad terén a zaplaviť územie aj za protipovodňovými hrádzami. Infiltrovaná voda z Dunaja potom prúdi ako podzemná voda smerom do Malého Dunaja, Mošonského ramena Dunaja a je drénovaná sieťou kanálov na Žitnom ostrove a na ostrove Szigetköz Obr. III.7.
Z geomorfologického hľadiska je dôležité, že pod žulovým skalným prahom, vystupujúcim z dna Dunaja pri Bratislave, kde začína náplavový kužeľ, je vysoký sklon Dunaja, s hodnotami 40 cm na km. Podobné andezitové a vápencové skalné dno je v úseku od Štúrova po Nagymaros. Toto spôsobuje zmenšenie sklonu Dunaja na menej ako 10 cm na km a tým aj ukončenie náplavového kužeľa Obr. III.8.
V Atlase SSR (Mazúr et al., 1980) Konček (in Mazúr et al., 1980, a in Mucha a Dub, 1966) definuje oblasť Podunajskej nížiny ako teplú, pri Bratislave mierne vlhkú (index zavlažovania Iz je menej ako –20), prechádzajúcu smerom na východ cez mierne suchú (Iz je –20 až 0), až do suchej (Iz je od 0 až do 60), s miernou zimou a s dlhším slnečným svitom.
Index zavlažovania podľa Končeka
Iz = R/2 + Δr – 10t – (30 – v2),
kde R – úhrn zrážok v mm
Δr – kladná odchýlka zrážok za december až február od 105 mm
t – priemerná teplota vzduchu v °C
v – priemerná rýchlosť vetra o 14 hod. v m/s
Počet letných dní s maximálnou teplotou nad 25 °C je viac ako 50 dní do roka (pri Bratislave viac ako 60 a v JV časti Žitného ostrova viac ako 70). Priemerná denná teplota pod 0 °C sa vyskytuje v oblasti Žitného ostrova 50 až 60 dní. Snehová pokrývka okolo 88 až 90 dní a jej maximálna hrúbka dosahuje v priemere 20 – 25 cm. Priemerné absolútne minimálne ročné teploty vzduchu sa pohybujú medzi –18 až –19 °C.
Trvanie slnečného svitu vo vegetačnom období prekračuje 1500 hod. (celoročne vyše 2000 hod.), čo je najdlhší slnečný svit na Slovensku. Začiatok žatvy raži ozimnej je pred 15. júlom (výsledok dĺžky slnečného svitu a teplôt). Pomerne vysoké teploty a veľká dĺžka slnečného svitu vo vegetačnom období je v podstate kladným rysom klímy, čo vytvára spolu s priestorovo variabilnou hĺbkou hladiny podzemnej vody, jej kolísaním zvlášť vhodné podmienky pre lužné lesy hlavne v záplavovom území ramennej sústavy Dunaja s osobitnou typickou skladbou biotopov. Z hľadiska poľnohospodárskej činnosti, je to najúrodnejšia oblasť, vyžadujúca povrchové závlahy, prípadne vysokú hladinu podzemných vôd.
Tarábek (in Mazúr at al., 1980) definuje klimaticko-geografický typ ako nížinná klíma s miernou inverziou teplôt, suchá až mierne suchá, teplá.
Suma priemerných denných teplôt (súčet) 10 °C a vyšších je 3000 – 3200 v roku. Priemerná mesačná teplota v januári je –1 až –4 °C, v júli 20,5 až 19,5 °C. Amplitúda priemerných mesačných teplôt je 22 až 24 °C (Obr. III.9). Ročný úhrn zrážok predstavuje 530 až 650 mm. Podunajská nížina ako celok je z hľadiska zrážok najsuchšou oblasťou na Slovensku. Premenlivosť ročných i mesačných úhrnov zrážok je vysoká (Obr. III.10), v extrémne suchých rokoch klesá k 300 mm a v mokrých rokoch stúpa k 1000 mm, V Podunajskej nížine býva do roka okolo 25 búrok. V zimných mesiacoch je výskyt búrok výnimočný.
Podunajská nížina ako celok je najteplejšou oblasťou Slovenska s rovnomerne rozdelenými teplotami. Najvyššia teplota sa dosahuje v júli, minimálna v januári. Počet dní, v ktorých teplota nevystúpi nad 0 °C je do roka v priemere 30.
V dlhodobom ročnom priemere prevláda severozápadný vietor a na druhom mieste podľa početností je vietor opačného smeru, teda juhovýchodný, čo prakticky znamená prevládanie vetra pozdĺž Dunaja, v oboch smeroch. Zastúpenie vetrov zo SZ je v lete väčšie ako v zime, v jeseni a v zime prevládajú o niečo viac juhovýchodné vetry. Celá oblasť je v údolí Dunaja pomerne veterná. Najveternejší je koniec zimy a začiatok jari, najpokojnejšia je jeseň.
Potenciálny výpar podľa Tomlaina (in Mazúr at al., 1980), sa pohybuje okolo 800 mm ročne, pri Bratislave menej, v spodnej časti Žitného ostrova viac. V období letného polroku je to nad 600 mm. Priemerný výpar z povrchu pôdy je 470 – 500 mm. Priemerný rozdiel medzi potenciálnym výparom a zrážkami v letom období je 340 – 345 mm.
Klimatické pomery sa z dlhodobého hľadiska zákonite menia (Tab. III.1). Holocénnu paleoklimatickú chronológiu a klimatickú chronológiu historickej doby popisuje napríklad Michalík at al. (1999). Od roku 800 do 1000 bola stabilná teplá klíma. Od roku 1000 do 1500 n.l. postupne poklesla priemerná teplota približne o 2 °C. Potom, približne od roku 1500 do roku 1870 bola pomerne nízka teplota, tzv. malá doba ľadová. Od roku 1870 teplota postupne stúpa avšak z ďaleka ešte nedosiahla teploty, ktoré boli pred a okolo roku 1000. V súčasnosti sa nachádzame už v druhej polovici interglaciálu, ktorý by mal skončiť o 16000 rokov novým zaľadnením. Vidíme, že v histórii ľudstva sa klimatické podmienky veľmi menili, od zaľadnení po teplú klímu tzv. klimatického optima. S tým súviselo aj kolísanie morských hladín, napríklad v holocéne (pred 12000 až 6000 rokmi) stúpla hladina mora o cca 120 - 130 m (v priemere 2 cm za rok!) a následne poklesla o cca 5 m. Ostatných niekoľko tisíc rokov (5000 – 6000) je morská hladina pomerne stabilná.
Lapin (1995), na základe pozorovaní od roku 1901 do 1990 konštatuje, že za ostatných 90 rokov priemerná ročná teplota vzduchu stúpla o 0,8 °C, potenciálna evapotranspirácia stúpla o 14 %, zrážkové úhrny poklesli o 15 %, poklesla relatívna vlhkosť vzduchu. Ak uvážime aj dlhodobý trend poklesu hladín podzemných vôd vrátane pôdnej vlhkosti, a tiež poklesu hladín a vodných plôch v ramenných sústavách pred rokom 1992, potom je možné konštatovať, že tieto zmeny signifikantne ovplyvnili prírodné prostredie, vrátane lokálneho poklesu vlhkosti a stúpnutia teplôt vzduchu.
Súčasné klimatické pomery oblasti Dunaja, závisia od cirkulačných pomerov vzduchu v Európe. Cirkulačné pomery v ovzduší sú prvoradým činiteľom ovplyvňujúcim počasie a klímu (Konček in Mucha V. Dub O., red., 1966). Počasie závisí predovšetkým od prúdenia vzduchu a prínosu vzduchových hmôt od morských a pevninských oblastí. Ročný výkyv teploty je v morskom vzduchu a povrchových vodách oceánu len 10 – 15 °C, kým v pevninskom vzduchu 50 – 60 °C. Podobne, rozdiely relatívnej vlhkosti vzduchu v pevninskom vzduchu medzi letom a zimou sú podstatne vyššie ako v morskom vzduchu. V oblasti Podunajskej nížiny sa najčastejšie nachádza vzduch pochádzajúci z miernych zemepisných šírok. Prenikajú sem však vzduchové hmoty arktického pôvodu a tropické vzduchové hmoty zo subtropických šírok. Ich striedaním vznikajú v našich oblastiach citeľné výkyvy teplôt a celkového charakteru počasia. V zimnom období, keď je rozdiel v teplotách medzi nízkymi a vyššími zemepisnými šírkami podstatne väčší ako v lete, aj prúdenie vzduchu je intenzívnejšie, a tým aj kolísanie teploty a charakteru počasia. Striedanie sa vzduchových hmôt a postup atmosferických frontov, na ktoré sa viaže najviac zrážok, riadi vývoj počasia. Zrážky, ktoré padajú v zime, padajú z morského vzduchu, ktorý preniká nad pevninu a vystupuje nad chladný pevninský vzduch. V lete pochádzajú zrážky hlavne z pevninského vzduchu. To znamená, že zimné zrážky nad pevninou súvisia s obehom vlahy medzi morom a pevninou, kým letné sú výsledkom hlavne obehu vlahy nad pevninou. Najnižšie teploty prinášajú vpády arktického vzduchu. Vtedy teplota silne klesá a obloha sa zvyčajne vyjasňuje. Na prúdenie vzduchu a zmeny teploty pôsobia mnohé ďalšie vplyvy. V Podunajskej nížine je to hlavne karpatské horstvo a vlastné údolie Dunaja.
Pre Podunajskú nížinu je typické časté striedanie sa prúdenia vzduchu a striedanie sa vzduchu rôzneho miesta vzniku, a to hlavne na jar. Na začiatku letného obdobia býva počasie nestále, so zväčšenou oblačnosťou a pomerne častými dažďami. V druhej polovici leta sa častejšie vyskytuje pekné počasie, pričom teploty vystupujú niekedy aj nad 35 °C. Pre začiatok jesene, v septembri a začiatkom októbra, je charakteristické jasné počasie, za ktorým obyčajne nasleduje zamračené počasie s častými zrážkami. V zimnom období bývajú v Podunajskej nížine, podobne ako v strednej Európe, najväčšie výkyvy teploty, a to v priebehu jednej zimnej sezóny ako aj v zimných obdobiach rôznych rokov. Trvalejší zimný ráz počasia býva len za prílevu studeného vzduchu z vyšších zemepisných šírok, a to hlavne z pevniny. Za takých podmienok sa vytvára trvalejšia snehová pokrývka a teploty klesajú v extrémnych prípadoch i pod – 30 °C. Normálna zima má však časté sneženie a častý odmäk.
Z hľadiska Dunaja a jeho ramien významnú úlohu majú aj ľadové úkazy. Priebeh ľadových úkazov a zamrznutia Dunaja v Bratislave uvádza Konček (in Mucha, Dub, 1966), Obr. III.11a. Ľadové úkazy sú nebezpečné aj z hľadiska tzv. ľadových povodní, ktoré vznikajú vytvorením ľadovej zápchy na toku. Príkladom takejto povodne je povodeň s kulmináciou 5. februára 1850, ktorá zaplavila starú Bratislavu, Obr. III.11b. Preto aj Zmluva z roku 1977 má jeden z cieľov „prevádzanie ľadov“.
Charakter pôdnych pomerov Podunajskej nížiny je určovaný vývojom klimatických podmienok, dlhodobými zmenami hladín podzemných vôd, zrážkami, zrnitostným zložením pôdy a sedimentov v zóne aerácie. Zloženie sedimentov od povrchu k hladine podzemnej vody modifikuje miestny vodný a vlhkostný režim aj pri rovnakej hĺbke hladiny podzemnej vody. Nezanedbateľnú úlohu má pritom technológia poľnohospodárskeho obrábania pôdy, prípravy pôdy pre vysádzanie lesa ako aj používanie závlah a simulovaných záplav. Riadenie vodného režimu je možné využiť ako opatrenie na zamedzenie vplyvu trendov dlhodobých i krátkodobých klimatických zmien.
Pôdy sa rozdeľujú na pôdne druhy a pôdne typy. Pôdne druhy sa ďalej delia na základe zrnitosti (textúra). Pôdne typy sa ďalej delia podľa charakteru pôdotvorného procesu v pôde.
Horniny, z ktorých vznikajú pôdy, sa zvetrávacími procesmi menia na zvetraliny určitej zrnitosti. Pôdu tvoria tieto zrniečka, zrnitostné prvky, (textúrne elementy) o rôznej veľkosti. Pôdne druhy rozlišujeme na základe zastúpenia jednotlivých zrnitostných frakcií s priemerom menším ako 2 mm na pôdy piesočné, hlinito-piesočnaté, piesčito-hlinité, hlinité, ílovito-hlinité, ílovité a íly. Podľa obsahu častíc nad 2 mm (skelet) rozdeľujeme pôdy na silno skeletnaté, skeletnaté a slabo skeletnaté (často používame aj iné prívlastky, napríklad štrkovité, kamenité a pod. Podľa obsahu ílu napríklad na ľahké, ťažké a pod., Obr. III.12.
Takéto rozdelenie má význam hlavne z hľadiska vlastností pôdy vzhľadom na schopnosť prijímať zrážky, prepúšťať zrážkovú vodu k hladine podzemnej vody a vzhľadom na kapilárne vlastnosti pôdy a zóny aerácie nasávať podzemnú vodu. S tým súvisí i technológia obrábania pôdy, závlahy, drenáž, a následne aj charakteristiky biotopov.
V oblasti pod Bratislavou sú pôdy podmienené transportom riečneho štrku, pieskov a plavenín. Sú to pôdy ľahké, prevážne piesočnaté s prímesou štrkov, smerom na JV hlinito-piesočnaté až hlinité, ktoré vznikli na nivných riečnych sedimentoch. Piesočnaté pôdy podmienené viatym pieskom sa nachádzajú od Palárikova smerom na Nové Zámky, Nesvady, Imeľ, Hurbanovo až k Moči. Okrem toho piesočnaté pôdy sú na viacerých lokalitách, napríklad pri Jelke, západne od Dunajskej Stredy, východne od Veľkého Medera a i.
Pôdne typy Obr. III.13 sú výsledkom pôdotvorného procesu za účinkovania špecifických pôdotvorných faktorov a podmienok na lokalite. Na území Podunajskej nížiny sú to predovšetkým rovinný terén riečnych náplavov Dunaja, špecifické klimatické podmienky s dlhým slnečným svitom, veľkým počtom teplých letných dní, zrážok je pomerne málo, ale na druhej strane sú vo vegetačnom období vysoké prietoky v Dunaji, občasné záplavy územia, a to v čase keď sa pôda tvorila a v časti územia je tomu tak aj dnes. Hĺbka hladiny podzemnej vody je rôzna, kolísanie hladiny podzemnej vody je pomerne veľké, s maximálnymi hladinami v letných mesiacoch.
V Podunajskej nížine nájdeme popri Dunaji a Malom Dunaji prevažne fluvizeme, nivné karbonátové pôdy na holocénnych aluviálnych sedimentoch. Profily týchto pôd majú obyčajne geologické zvrstvenie, na vrchu sú obyčajne hliny, pod nimi štrková vrstva, potom piesčitá a zase štrková. Vlastnosti týchto pôd sú závislé od zrnitostného a chemického zloženia sedimentov, režimu podzemných a povodňových vôd. Charakteristické je veľké kolísanie hladiny podzemnej vody spôsobené hlavne režimom kolísania prietokov vody v Dunaji. Človek výrazne ovplyvnil vývoj pôdy budovaním hrádzí a ovplyvňovaním režimu podzemných a povrchových vôd. Väčšina našich fluvizemí sa prestala zaplavovať povodňami a začínajú sa postupne premieňať na terestrické pôdy. Podmáčané fluvizeme sa menia na glejové pôdy.
Na starších riečnych hlinách a povodňových kalových usadeninách s nehlboko ležiacim štrkovým povrchom a hladinou podzemnej vody v štrkoch (alebo vo všeobecnosti v hlbších polohách) sa vytvorili karbonátové micelárne černozeme obsahujúce v humusovom horizonte vyzrážaný uhličitan vápenatý (od Podunajských Biskupíc smerom na Rastice, Šamorín a Dunajskú Stredu). Tieto sa vytvorili hlavne v dôsledku malých zrážok a vyššieho obsahu uhličitanu vápenatého v povodňových hlinách a sedimentoch. Smerom do vlhších území je táto černozem viac vylúhovaná a prechádza smerom k hnedozemnému typu.
Na aluviálnych náplavoch s vysokou hladinou podzemnej vody, pravidelne zaplavovaných a na podmáčaných sprašiach sa vytvorili lužné pôdy kvalitou blížiace sa černozemi (južne, východne a severne od Dunajskej Stredy smerom k Dunaju a Malému Dunaju). Lužná pôda vznikla na aluviálnej nive s obsahom karbonátovej zložky a s vplyvom mineralizovanej (kalcium bikarbonátovej) podzemnej vody s vyššou hladinou. Pôvodnú vegetáciu tvorili hlavne hydrofilné spoločenstvá. Hlavným pôdotvorným procesom tu bolo výrazné a hlboké hromadenie kvalitných humusových látok v podmienkach zvýšeného prevlhčenia pôdy z minerálne bohatých podzemných vôd (350 – 1000 mg/l). V miestach, kde je hladina podzemnej vody stále blízko pod terénom (okolo 0,5 m), sa vytvorili glejové lužné pôdy, podobné černozemi. Časť dnešných lužných pôd vznikla z glejových pôd po znížení hladiny podzemných vôd. Na holocénnych agradačných valoch, kde je hladina podzemnej vody mierne hlbšie, sa vytvorili lužné černozeme.
Glejové pôdy sa tvoria na aluviálnych sedimentoch nív v terénnych depresiách, kde je hladina podzemnej vody blízko k povrchu. Pôvodnú vegetáciu tvorili rôzne močiarne a vlhkomilné spoločenstvá. Hlavným pôdotvorným procesom je glejový proces v redukčnom prostredí, pri ktorom dochádza predovšetkým k redukcii najmä železitých a manganičitých zlúčenín za účasti anaerobných mikroorganizmov.
V Podunajskej nížine sa vytvorili v terénnych depresiách a mŕtvych ramenách rašeliny a rašelinové pôdy (napr. Pusté Uľany, Jurský Šúr, Dunajská Streda, Veľký Meder). Smerom na Komárno sa zase vytvorili čiastočne zasolené pôdy (medzi Komárnom a Veľkým Mederom, pri Dunajskej Strede, pri Komárne).
Z hľadiska inundačného územia spomenieme ešte surovú fluvizem, nivnú pôdu (rambla), ktorá je veľmi mladou riečnou uloženinou alebo i oderodovanou plochou, na ktorej povrchu ešte nie je viditeľný humusový horizont. Ide o pôdu ľahkú, piesčitú, často štrkovitú. Takéto pôdy sú dôležité hlavne z hľadiska prirodzeného vývoja a uchytenia pre inundáciu typických druhov porastu, hlavne obnova drevín zo semena, najmä domácich vŕb a topoľov (asociácie Salici - Populetum), ktorá sa deje výlučne na takýchto pôdach. Na ílovitých, hlinitých a jemno piesočnatých substrátoch sa uchytáva vŕba biela a topoľ biely i sivý, kým topoľ čierny sa uchytáva len na štrkoch (Šomšák et al., 2002).
III.4.3 Klimatické zmeny, podzemné vody a poľnohospodárska činnosť
Vzhľadom na očakávané klimatické zmeny konštatujeme, že z hľadiska poľnohospodárskej výroby ide hlavne o zmeny teploty, ktoré vyvolávajú zmeny evapotranspirácie a v nej predovšetkým zmeny v transpiračných procesoch rastlín. Takáč (2001), na základe výsledkov výpočtov scenárov klimatických zmien, konštatuje, že postupne, v jednotlivých časových horizontoch, príde k zvyšovaniu teploty a k znižovaniu počtu dní so snehovou pokrývkou pod 20 dní. V priebehu zimy bude dochádzať častejšie k topeniu snehu a k rozmŕzaniu povrchovej vrstvy pôd, čo umožní infiltráciu snehovej vody ešte v zimných mesiacoch. Následne, v marci, infiltrácia poklesne, pretože snehová pokrývka bude roztopená už skôr. V dôsledku zvyšovania teploty dôjde k zvyšovaniu priemerných úhrnov potenciálnej evapotranspirácie (výparnosti). Hodnota priemerného obsahu vody v hĺbke do 100 cm sa zmenší. Ročné úhrny zrážok, ani ich rozloženie počas vegetačného obdobia, nebudú postačujúce na pokrytie vlahovej potreby plodín, preto pre poľnohospodársku výrobu bude nevyhnutná doplnková závlaha. Skutočná evapotranspirácia a podiel výparu z pôdy a transpirácie rastlinami bude závisieť od hĺbky hladiny podzemnej vody, od technológie poľnohospodárskej výroby a hlavne od závlah. Očakáva sa, že zvýšenie závlahových množstiev bude potrebné koncom mája a v letných mesiacoch júl a august.
Z hľadiska očakávaných klimatických zmien je možné pomocou zariadení vybudovaných v rámci výstavby vodného diela Gabčíkovo, prípadne ďaľších opatrení, ovplyvniť vodný režim. Čo sa týka vodného režimu bude potrebné od marca zvýšiť hladiny podzemných vôd, čo sa na niektorých plochách prejaví zníženou potrebou závlah. Okrem toho bude treba zvýšiť hladiny vody v ramenných sústavách a v kanálovej sieti Žitného ostrova a ostrova Szigetköz. Zvýšenie hladín vody v starom koryte Dunaja pretekanými prehrádzkami podobnými ako pri Dunakiliti je možné podstatne znížiť drénovanie podzemnej vody starým korytom Dunaja. V ľavostrannej i pravostrannej ramennej sústave sa odporúča častejšie realizovať simulované záplavy.
Dunaj pod Bratislavou bol v minulosti voľne tečúcou, divočiacou, širokou a meandrujúcou riekou s nespočetnými veľkými a malými ramenami a so širokou inundáciou siahajúcou ďaleko za dnešné protipovodňové hrádze. Malý Dunaj a Mošonský Dunaj boli v minulosti hlavnými ramenami-korytami Dunaja, využívanými aj na plavbu. Inundácia, teda pravidelne zaplavované územie s meandrujúcimi ramenami Dunaja, sa rozkladala minimálne od Malého Dunaja na severe až za Mošonský Dunaj na juhu. Hĺbky vody v korytách Dunaja a rýchlosti prúdenia vody boli podstatne menšie, voda bola viac rozdelená do riečnych ramien, hladina sa počas povodní viac rozlievala po teréne a hladina podzemnej vody bola podstatne plytšie pod terénom. V tom čase, na náplavovom kuželi pod Bratislavou, kde sa Dunaj rozvetvoval, sa usadzovali hrubé štrky a piesky prinášané Dunajom z územia nad Devínskou bránou a náplavový kužeľ postupne rástol.
Za veľkých vôd si Dunaj prekladal svoje koryto a dochádzalo k rozsiahlym záplavám. Delenie vody nevyhovovalo ani plavbe. Preto sa už v 13. storočí stavali ochranné hrádze, hlavne na pravej strane Dunaja, a hlavné koryto Dunaja sa začalo postupne napriamovať. Vzhľadom na zlepšenie plavebných podmienok sa v roku 1886 prikročilo k regulácii toku na strednú vodu (rkm 1880 - 1747). Práce boli ukončené v roku 1896, ale nesplnili očakávanie hlavne pre plavbu počas nízkych vodných stavov. Preto sa začiatkom 20. storočia pristúpilo k úprave Dunaja na tzv. malú vodu. Čiastočným uzatvorením ramien sa predovšetkým sústredil hlavný tok do jedného ramena, prúdenie vody sa ustálilo, pre zabezpečenie plavby za nízkych vodných stavov sa vybudovali výhony a nebezpečné brody sa bagrovali.
K ďalším zmenám vodného režimu došlo po rozsiahlych úpravách hlavného koryta rieky, spojených s plavbou a s protipovodňovou ochranou, v 1970-tych rokoch. Bagrovanie štrku z dna Dunaja na slovenskej i maďarskej strane malo za následok pokles hladiny vody v rieke, a aj hladín podzemných vôd. Tendencia klesajúcej hladiny vody bola neustále udržiavaná pokračujúcou eróziou dna rieky aj pričinením vodných stavieb na rakúskom a nemeckom úseku Dunaja a bagrovaním Dunaja nad Bratislavou.
Prietoky v Dunaji, výška dna Dunaja a samozrejme i úprava toku a brehov, boli pre inundačné územie pred uvedením vodného diela Gabčíkovo rozhodujúce. Od prietokov v Dunaji závisel nielen stav hladiny vody v Dunaji a tým aj splavnosť Dunaja, ale aj prietoky a hladiny vody v ramennej sústave a stav hladín podzemných vôd.
Pred úplným uzatvorením ramien a pred oddelením ramennej sústavy od Dunaja, v rokoch 1955 až 1961, teda ešte pred sústredením vody do hlavného koryta Dunaja kvôli plavbe, ale už po napriamení Dunaja a postavení protipovodňových hrádzí, voda tiekla ramenami na maďarskej a slovenskej strane aj pri nízkych prietokoch v Bratislave. Zachovali sa nám o tom záznamy v publikácii Mucha, Dub (1966). Tieto záznamy uvádzame v Tabuľke III.2.
Výstavbou protipovodňových hrádzí za oboma brehmi Dunaja a opevnením jeho brehov sa obmedzilo jeho pôvodné voľné meandrovanie a takto napriamené a upravené hlavné koryto, sme pred rokom 1992 poznali ako Dunaj (Obr. III.14).
Pred opevnením a zvýšením brehov Dunaja a pred uzavretím vtokov ramien (v prvej polovici 20. storočia) voda tiekla v ramenách počas všetkých prietokov v Dunaji, na niektorých miestach vtekala, inde vytekala, a to na maďarskej aj na slovenskej strane. V hlavných prietočných ramenách nebola ani občasne stojatá voda.
Tab. III.2 Percentá prietoku Dunaja a v zátvorke jeho ramien spolu s inundačným územím
pre rôzne prietoky v Dunaji v Bratislave Propeleri (Mucha, Dub, 1966)
|
Dátum |
17.1.1961 |
21.9.1960 |
20.-22.6.1961 |
16.-17.5.1961 |
13.-14.7.1955 |
|
Prietok |
1005 m3/s |
1958 m3/s |
2998m3/s |
4002m3/s |
6702m3/s |
|
rkm |
% |
||||
|
1869,04 Bratislava |
100 (0) |
100 (0) |
100 (0) |
100 (0) |
100 (0) |
|
1860,00 |
92 (8) |
89 (11) |
92 (8) |
92 (8) |
65 (35) |
|
1851,63 |
79 (21) |
81 (19) |
88 (12) |
89 (11) |
87 (13) |
|
1847,85 |
84 (16) |
95 (5) |
85 (15) |
70 (30) |
58 (42) |
|
1842,40 |
99 (1) |
97 (3) |
83 (17) |
68 (22) |
50 (50) |
|
1833,10 |
76 (24) |
74 (26) |
62 (38) |
65 (35) |
48 (52) |
|
1825,00 |
88 (12) |
96 (4) |
78 (22) |
67 (33) |
53 (47) |
|
1821,07 |
65 (35) |
82 (18) |
66 (34) |
51 (49) |
40 (60) |
|
1816,85 |
84 (16) |
84 (16) |
76 (24) |
64 (36) |
50 (50) |
|
1810,40 |
99 (1) |
85 (15) |
68 (32) |
63 (37) |
43 (57) |
|
1806,00 |
99 (1) |
99 (1) |
89 (11) |
92 (8) |
73 (27) |
|
1802,37 |
99 (1) |
99 (1) |
90 (10) |
99 (1) |
75 (25) |
% v Dunaji (v ramenách, pri prietokoch nad 2000 m3/s postupne aj s prietokmi inundáciou po vybrežení).
Pri prietokoch nad 4500 m3/s bola už zaplavená celá inundácia.
Od toho času je pôvodne trvalé prepojenie Dunaja s jeho ramenami obmedzené len na obdobie, kedy v Dunaji tečie viac vody (2500 – 4000 m3/s) ako je priemerný prietok (2025 m3/s). Tak sa stalo, že pred prehradením Dunaja v roku 1992 voda v takmer všetkých dunajských ramenách tiekla v priemere len 17 dní do roka (CEC, 1992).
Režim prietokov v ramenách bol závislý vždy od prietokov v Dunaji a bol ovplyvňovaný stavom opevnenia brehov hlavného koryta Dunaja a výškou brehových priepadov a vtokov do pôvodných ramien ako aj úrovňou dna Dunaja.
Podobne, hladina podzemnej vody, hlavne v pririečnej zóne, ale aj v širšom území, závisela vždy od hladiny vody v Dunaji a bola ovplyvnená hladinou vody v ramennej sústave i v kanálovej sústave za ochrannými protipovodňovými hrádzami. Typické prietoky v Dunaji v tomto období sú uvedené v Tab. III.3.
Tab. III.3 Typické prietoky v Dunaji v Bratislave (Bratislava, 1901 - 1950), Cec (1992)
|
Charakteristika prietoku |
prietok (m3/s) |
|
Priemerný prietok |
2025 |
|
Mesiac s minimálnym priemerným prietokom |
1441 (december) |
|
Mesiac s maximálnym priemerným prietokom |
2785 (jún) |
|
Typický nízky prietok (priemer ročných miním) |
848 |
|
Typický vysoký prietok (priemer ročných maxím) |
5316 |
Pomer medzi priemerným maximálnym mesačným letným (jún) a priemerným minimálnym mesačným zimným (december) prietokom v Dunaji je 1,93, čo znamená, že prietok v lete je v priemere približne dvojnásobkom zimného prietoku, Tab. III.3.
Výška hladiny vody v pozdĺžnom profile Dunaja na základe meraní z rokov 1969 – 1976 (Spoločný zmluvný projekt 1976) je pre prietok Dunaja 2300 m3/s uvedená na Obr. III.8.
Pred uvedením vodného diela do prevádzky (október 1992), podľa stavu koryta Dunaja aký bol v roku 1980, skoro vo všetkých ramenách tiekla voda až pri prietokoch v Dunaji vyšších ako 3500 m3/s, čo predstavovalo asi 17 dní do roka. V niektorých hlavnejších ramenách tiekla voda pri prietokoch nad 2500 m3/s, čo predstavovalo približne 3 mesiace do roka. Stav pred prehradením v roku 1992 bol zrejme ešte o niečo horší, ako je opísaný v správe CEC (1992).
Typické vysoké prietoky boli v lete, hlavne v júni a júli, kedy pred prehradením tiekla voda zvyčajne vo väčších ramenách.
Typické nízke prietoky boli v októbri, novembri, decembri, kedy v ramenách voda obyčajne netiekla, v časti ramien bola stojatá voda a časť ramien bola suchá. Typická prietoková situácia stavu z roku 1980 je v Tab. III.4.
Tab. III.4 Typické prietokové situácie v ramennej sústave pred prehradením (Cec, 1992)
|
Prietoková situácia |
Prietokové podmienky v r. 1980 (m3/s) |
Hladina vody v Dunaremete (m) |
Rýchlosti prúdenia v Dunaji v Dunaremete (m/s) |
Priemerná dĺžka trvania (deň do roka) |
Frekvencia (jav/rok) |
|
Prúdenie v Dunaji viazané na dno medzi výhonmi |
<1000 |
2,3 |
1000 m3/s => 1,4 m/s |
< 1000 m3/s, 13 dní |
niekoľko krát |
|
Prúdenie v hlavnom koryte a trvalých ramenách |
<1800 |
3,7 |
1800 m3/s => 1,8 m/s |
1000-1800 m3/s, 42 dní |
niekoľko krát |
|
Prúdenie v máloktorých ramenách |
1800-2500 |
3,7-4,5 |
1,8-2,0 m/s |
1800-2500 m3/s, 122 dní |
niekoľko krát |
|
Prúdenie v niektorých ramenách |
2500-3500 |
4,5-5,2 |
2,0-2,2 m/s |
2500-3500 m3/s, 78 dní |
niekoľko krát |
|
Prúdenie v skoro všetkých ramenách |
3500-4500 |
5,2-5,6 |
2,2-2,3 m/s |
3500-4500 m3/s, 17 dní |
niekoľko krát |
|
Úplné zaplavenie inundácie |
> 4500 |
5,6 |
4500 m3/s => 2,3 m/s |
> 4500 m3/s, 4 dni |
raz za rok |
|
Hlboké zaplavenie inundácie |
6000 |
6,2 |
6000 m3/s => 2,4 m/s |
>6000 m3/s < 1 deň |
raz za 3-4 roky |
Predchádzajúce ľudské činnosti viedli k nasledujúcim dlhodobým a postupným zmenám:
Zamedzeniu meandrovania Dunaja a tým aj skrátenie jeho toku a zvýšenie sklonu rieky.
Podstatnému zväčšeniu hĺbky vody a rýchlosti prúdenia v hlavnom koryte Dunaja, zvýšeniu erózie dna, zníženiu sedimentácie riekou prinášaných štrkov a pieskov.
Podstatnému zníženiu prínosu piesku a štrku cez Devínsku bránu do priestoru Dunaja pod Bratislavou.
Podstatnému zmenšeniu rýchlosti prúdenia v riečnych ramenách, prerušovaniu prietoku v ramenách, usadzovaniu jemnozrnných sedimentov a zazemňovaniu ramien namiesto pôvodnej erózie a sedimentácie hrubších piesčitých a štrkovitých frakcií, postupnému zamedzovaniu meandrovania hlavného toku Dunaja aj jeho ramien.
Odpojeniu ramien od hlavného toku, vysýchaniu mnohých ramien.
Všeobecnému a postupnému poklesu dna a hladín vody v Dunaji po celej jeho dĺžke od Bratislavy až po Budapešť.
Všeobecnému a postupnému poklesu hladín podzemných vôd a zmenám smerov prúdenia podzemných vôd.
Vysýchaniu mŕtvych ramien a ramien za protipovodňovými hrádzami, ich postupnému zazemňovaniu a zavážaniu odpadmi.
Dlhodobý priebeh prietokov v Dunaji v profile v Bratislave a dlhodobý pokles hladín vody v Dunaji v Bratislave, Rusovciach, Medveďove a Komárne sú zjavné z dát nameraných Slovenským hydrometeorologickým ústavom, zobrazených na Obr. III.17.
Výška hladiny podzemných vôd bola meraná na slovenskom i maďarskom území na viac ako 1000 pozorovacích vrtoch. Príkladom môže byť mapa hĺbok hladín podzemných vôd na maďarskom území (Scharek, 1991), Obr. III.18. Na mape je možné vidieť hlboko poklesnutú hladinu podzemných vôd v hornej časti územia pri Rajke. Podobne, na Obr. V.81 sú vynesené pozorovacie objekty hladín podzemných vôd na slovenskom území.
Dlhodobý pokles hladín podzemných vôd je evidentný na väčšine územia (FNSCU, 1995). Tento pokles je tiež možné vidieť na hydrografoch pozorovacích vrtov, Obr. III.19, ktoré sú nakreslené ako závislosť kolísania hladín podzemných vôd v závislosti na čase, Obr. III.20a, Obr. III.21a, Obr. III.22a a Obr. III.23a (slovenské územie) a Obr. III.20b, Obr. III. 21b, Obr. III. 22b a Obr. III. 23b (maďarské územie). Na základe referenčnej hladiny podzemnej vody v roku 1962 a 1992 (FNSCU, 1995) bola zostavená mapa vyjadrujúca rozdiel hladiny podzemných vôd medzi rokmi 1962 a 1992, to znamená obdobie 30 rokov pred uvedením vodného diela Gabčíkova do prevádzky, Obr. III.24. Veľký pokles hladín podzemných vôd za posledných 30 rokov pred uvedením vodného diela do činnosti je zreteľný hlavne v hornej časti Žitného ostrova, pod Bratislavou. Poklesy hladín podzemných vôd sú spôsobené hlavne poklesom dna a hladín v Dunaji, a tiež skrátením toku (odrezané meandre), obmedzovaním prúdenia v ramennej sústave v inundácii a v ramenách Dunaja vrátane Malého Dunaja a Mošonského ramena Dunaja. Pokles hladiny podzemnej vody v predchádzajúcom 30 ročnom období pred uvedením Vodného diela Gabčíkovo do prevádzky negatívne ovplyvnil prírodné podmienky hlavne v inundácii a vo všeobecnosti negatívne ovplyvnil poľnohospodárstvo, lesné hospodárstvo a tiež zásoby podzemnej vody. Negatívne ovplyvnenie prírodného prostredia je popísané v mnohých publikáciách, čitateľa odkazujeme napríklad na publikáciu (FNSCU, 1995, 1999 str. 15 a ďalšie články v publikácii) a publikáciu Mucha at al., 1999.
Kvalitu povrchovej a podzemnej vody z rokov 1954 až 1961 poznáme z práce Antonič a Jacko (1966) „Hydrochémia československého úseku Dunaja a jeho prítokov“. Priemerné hodnoty vybraných ukazovateľov chemického zloženia vôd Dunaja v Bratislave, v rkm 1871, za roky 1954 – 1961 sú uvedené v Tab. III.5.
Tab. III.5 Priemerné hodnoty vybraných ukazovateľov chemického zloženia vôd Dunaja
v Bratislave (rkm 1871) za roky 1954 – 1961
Ukazovateľ |
Množstvo |
Ukazovateľ |
Množstvo |
|
Teplota vody [oC] |
10,7 |
HCO3- [mg/l] |
182,8 |
|
Merná vodivosť (ECT) [mS/m] |
29,6 |
Fe3+ [mg/l] |
0,13 |
|
pH [-] |
7,6 |
Ca2+ [mg/l] |
51,9 |
|
CHSKMn [mgO2/l] |
5,1 |
Mg2+ [mg/l] |
12,6 |
|
BSK5 [mgO2/l] |
2,2 |
Na+ [mg/l] |
7,1 |
|
O2 [mg/l] |
9,7 |
K+ [mg/l] |
4,4 |
|
NO2 [mg/l] |
0,10 |
NH4+ [mg/l] |
0,28 |
|
NO3- [mg/l] |
3,7 |
Mn2+ [mg/l] |
0 |
|
PO43- [mg/l] |
0,10 |
Fenoly [mg/l] |
0,019 |
|
SO42- [mg/l] |
31,6 |
B [ng/l] |
27-54 |
|
Cl- [mg/l] |
12,4 |
|
|
Podľa vtedajších kritérií autori Antonič a Jacko hodnotili kvalitu vôd Dunaja za vyrovnaných a klesajúcich prietokov ako vodu prípustnú III. triedy čistoty. BSK5 sa pohybovalo okolo 2,5 mg O2/l a oxidovateľnosť podľa Kubela (CHSKMn) 4,4 – 7,1 mg O2/l. Zdroje znečistenia, ako odpadové vody z Bratislavy a znečistenie prítokov, sa prejavovali lokálne v úzkych pásoch pri brehu. Čistota vody sa výrazne znižovala v dobe cukrovarníckej kampane, napríklad pod vtokom rieky Moravy. Nárazové znečistenie vplyvom prírodných podmienok (vysoké prietoky) sa prejavovali zvýšením hodnôt BSK5 na okolo 3,5 mg O2/l a oxidovateľnosti podľa Kubela nad 6,0 mg O2/l.
Z hľadiska saprobiológie československého úseku Dunaja Rothschein a Hanzlíková (1966) definujú československý úsek Dunaja ako β-mezosapróbnu rieku. Autori píšu, že znečistenie Dunaja minerálnymi olejmi pod Bratislavou sa v posledných rokoch zvýšilo a je najväčším nebezpečenstvom pre Dunaj. Oleje vytvárajú povlaky na povrchu vody, zaolejovávajú brehy. Ostatné znečistenie predstavujú odpadové vody z Viedne, ktoré v množstve cca 3 m3/s odchádzajú bez čistenia do Dunaja a odpadové vody z Bratislavy.
Priemerný stav čistoty dunajskej vody, indikovaný jeho oživením, je pri vstupe na slovenské územie prakticky rovnaký, ako v mieste, kde Dunaj opúšťa slovenské územie.
Antonič a Jacko (1966) ďalej píšu, že vážnym problémom sa stáva znečistenie Dunaja olejmi následkom výstavby veľkých rafinérií v Bratislave a Almásfüzitö. Minerálne oleje znečisťujú prevažne brehy Dunaja. Tým sú ohrozené aj možnosti vodohospodárskeho využitia Dunaja a bude treba urobiť energické kroky na zneškodnenie tohoto závažného znečistenia.
Čo sa týka podzemných vôd Antonič a Jacko (1966) píšu, že by v priestore južne od Bratislavy predpokladali približne také zloženie ako má dunajská voda. Očakávali by množstvo rozpustných látok 250 až 300 mg/l, obsah síranov 30 – 50 mg/l, chloridov 10 – 15 mg/l, dusičnanov 3-5 mg/l a železo len v stopových množstvách. Pri optimálnych filtračných vlastnostiach štrkopieskového zvodneného prostredia by vody nemali obsahovať produkty rozkladu organických látok (dusitany, amoniak, fosforečnany), prípadne mikrobiologické oživenie. Lenže, rozbormi sa zistilo na celom území pod Bratislavou ovplyvnenie odpadovými vodami. Napríklad v oblasti Prievozu, Podunajských Biskupíc, Mosta na Ostrove, Vrakuni a smerom k Ivánke pri Dunaji je v podzemných vodách okolo 800 – 2200 mg/l rozpustených látok, obsah síranov okolo 180 – 870 mg/l, chloridov 40 – 50 mg/l, dusičnanov 15 – 170 mg/l, vápnika 160 – 250 mg/l, horčíka 40 – 50 mg/l, sodíka do 90 mg/l, draslíka do 10 mg/l a železa do 1,3 mg/l. Tieto extrémne hodnoty však nie sú rovnomerne rozdelené, ale sa z miesta na miesto menia. Sprievodným znakom znečistenia podzemných vôd je aj mikrobiologická závadnosť (baktériami z čeľade Enterobacteriaceae), najmä tam, kde prevláda infiltrácia z Dunaja. Zloženie sa pohybuje v rámci nasledujúcich hodnôt: sírany 40 – 80 mg/l, chloridy 15 – 30 mg/l, dusičnany 1 – 10 mg/l. Zistili sa aj vysoké výskyty dusičnanov aj do 100 mg/l, čo autori v Podunajskej nížine nepovažujú za ojedinelý jav.
Smerom do územia Žitného ostrova, oblasť medzi Šamorínom, Tomášovom, až Gabčíkovom, Dunajskou Stredou a Jahodnou sa pohybuje celková mineralizácia do 700 mg/l, sírany obyčajne do 100 mg/l a vyššie hodnoty sú smerom k Malému Dunaju. Podobne smerom k Malému Dunaju stúpa aj obsah chloridov a dusičnanov, pri Malom Dunaji aj nad 100 mg/l.
Od tej doby sa veľa zmenilo, aspoň v úseku Vodného diela Gabčíkovo a nad Bratislavou. Predovšetkým, vo Viedni postavili čistiacu stanicu komunálnych odpadových vôd, čistiacu stanicu rafinérie Schwechat, podobne v Bratislave v rámci výstavby Petržalky a Sústavy vodných diel postavili čistiacu stanicu odpadových vôd v Petržalke a v Slovnafte. Boli postavené cesty, vodovody (10), kanalizácie (28) a čistiarne (14) odpadových vôd a nové vodné zdroje v obciach Žitného ostrova. Zmenili sa chemické prípravky používané v poľnohospodárstve. Bol vybudovaný monitoring prírodného prostredia na oboch stranách Dunaja (napr. Scharek, ed., 1991)
Výsledky monitorovania, uvádzané v nasledujúcich kapitolách poukážu na dnešný stav prírodného prostredia a kvalitu povrchových a podzemných vôd. Ako príklad toho čo sa dnes monitoruje a aké sú normy na pitnú vodu, môžu poslúžiť vodné zdroje Šamorín a Rusovce, Tab. III.6.
Tab. III.6 Príklad monitorovaných ukazovateľov kvality podzemných vôd pre vodné zdroje
(vodný zdroj Šamorín dňa 6.12.1999 a vodný zdroj Rusovce dňa 17.10.2000)
|
Ukazovateľ |
Jednotka |
Vyhláška MZ |
Slovenská norma |
Lokalita |
|
|
151/2004 |
STN 757111 |
Šamorín |
Rusovce |
||
|
teplota vody |
°C |
8-12 |
12 |
11 |
11,4 |
|
vzhľad po vzorkovaní |
|
|
|
číra |
|
|
zákal |
ZF |
5 |
|
bez |
0,90 |
|
farba |
mg/l |
20 |
|
bez |
|
|
pach |
|
bez pachu |
|
bez |
0 |
|
vodivosť |
mS/m |
125 |
100 |
36,6 |
46,0 |
|
reakcia vody – pH |
|
6,5-8,5 |
8,5 |
7,70 |
7,32 |
|
ZNK8,3 / Acidita pH 8,3 |
mmol/l |
|
|
0,12 |
|
|
KNK4,5 / Alkalinita pH 4,5 |
mmol/l |
|
|
3,9 |
3,700 |
|
CHSKMn |
mg/l |
3 |
3 |
0.8 |
0,600 |
|
rozpustené látky-sušené |
mg/l |
1000 |
1000 |
261 |
303,0 |
|
vápnik a horčík |
mmol/l |
1,1-5 |
5 |
2,2 |
2,200 |
|
vápnik |
mg/l |
>30 |
>30 |
66,1 |
62,12 |
|
horčík |
mg/l |
10-30 |
30 |
13,4 |
15,81 |
|
sodík |
mg/l |
200 |
|
10,9 |
10,63 |
|
draslík |
mg/l |
|
|
2,6 |
1,426 |
|
železo |
mg/l |
0,2 |
0,3 |
<0,01 |
0,001 |
|
mangán |
mg/l |
0,05 |
0,1 |
<0,05 |
0,014 |
|
amónne ióny |
mg/l |
0,5 |
0,5 |
<0,05 |
<0,030 |
|
dusitany |
mg/l |
0,1 |
0,1 |
<0,01 |
<0,030 |
|
dusičnany |
mg/l |
50 |
50 |
1,50 |
1,42 |
|
chloridy |
mg/l |
100 |
100 |
18,6 |
17,6 |
|
sírany |
mg/l |
250 |
250 |
23,5 |
32,2 |
|
fosforečnany |
mg/l |
|
|
<0,01 |
<0,100 |
|
kremičitany |
mg/l |
|
|
5,1 |
|
|
hydrogénuhličitany |
mg/l |
|
|
238 |
226 |
|
fluoridy |
mg/l |
1,5 |
1,5 |
<0,1 |
<0,2 |
|
tenzidy aniónové |
mg/l |
|
0,2 |
<0,05 |
<0,04 |
|
fenoly - prchavé s vodnou parou |
mg/l |
|
0,05 |
<0,03 |
<0,005 |
|
arzén |
μg/l |
10 |
10 |
<5 |
<1,2 |
|
kadmium |
μg/l |
3 |
3 |
<0,5 |
<0,04 |
|
olovo |
μg/l |
10 |
10 |
< 1 |
0,2 |
|
ortuť |
μg/l |
1 |
1 |
<0,36 |
<0,05 |
|
hliník |
mg/l |
0,2 |
0,2 |
<0,03 |
0,028 |
|
meď |
μg/l |
1000 |
500 |
3 |
7,58 |
|
zinok |
mg/l |
3 |
3 |
<0,02 |
0,103 |
|
chróm |
μg/l |
50 |
50 |
<1 |
0,19 |
|
nikel |
μg/l |
20 |
20 |
<1 |
1,69 |
|
antimón |
μg/l |
5 |
5 |
<3 |
<0,52 |
|
absorbancia |
|
0,08 |
0,08 |
0,01 |
<0,01 |
|
látky extrahovateľné nepolárne - UV |
mg/l |
|
0,05 |
<0,01 |
|
|
nepolárne extrahovateľné látky - IR |
mg/l |
|
|
0,03 |
<0,03 |
|
TOC |
mg/l |
5 |
|
2,03 |
1,261 |
|
fluorantén |
ng/l |
|
|
<3 |
<5 |
|
polycyklické aromatické uhľovodíky |
ng/l |
100 |
200 |
<30 |
|
|
benzo(b)fluorantén |
ng/l |
|
|
<3 |
<5 |
|
benzo(k)fluorantén |
ng/l |
|
|
<3 |
<5 |
|
benzo(g,h,i)perylén |
ng/l |
|
|
<3 |
<5 |
|
indeno(1,2,3-c,d)pyrén |
ng/l |
|
|
<30 |
<5 |
|
benzo(a)pyrén |
ng/l |
10 |
|
<3 |
<1 |
|
chloroform |
μg/l |
40 |
40 |
<0,01 |
0,111 |
|
tetrachlórmetán |
μg/l |
2 |
2 |
<0,01 |
<0,02 |
|
1,2 dichlóretán |
μg/l |
3 |
3 |
<0,04 |
<0,1 |
|
1,1,2,2 tetrachlóretén |
μg/l |
10 |
40 |
<0,01 |
<0,5 |
|
1,1,2 trichlóretén |
μg/l |
10 |
70 |
<0,01 |
<0,1 |
|
benzén |
μg/l |
1 |
1 |
<1 |
0,036 |
|
dichlórbenzény |
μg/l |
0,3 |
0,3 |
<0,01 |
<0,01 |
|
chlórbenzén |
μg/l |
10 |
10 |
<0,001 |
<0,020 |
|
dichlórfenoly |
μg/l |
2 |
2 |
<0,001 |
|
|
dichlórdifenyltrichloretán |
μg/l |
|
|
<0,005 |
|
|
hexachlórbenzén |
ng/l |
|
|
<2 |
<10 |
|
heptachlór |
ng/l |
|
|
<2 |
<10 |
|
chlóretén |
μg/l |
|
0,5 |
<0,05 |
|
|
lindan |
μg/l |
|
|
<0,002 |
<0,01 |
|
metoxychlór |
μg/l |
|
|
<0,002 |
<0,05 |
|
toluén |
μg/l |
50 |
50 |
<5 |
0,033 |
|
xylén |
μg/l |
100 |
100 |
<10 |
<0,02 |
|
termotolerantné koliformné baktérie |
KTJ/100 ml |
|
0 |
0 |
0 |
|
koliformné baktérie |
KTJ/100 ml |
0 |
0 |
0 |
0 |
|
fekálne streptokoky |
KTJ/100 ml |
0 |
0 |
0 |
0 |
|
mezofilné baktérie |
KTJ/1 ml |
20 |
100 |
0 |
1 |
|
psychrofilné baktérie |
KTJ/1 ml |
200 |
500 |
0 |
0 |
|
železité baktérie – kultivačne |
KTJ/100 ml |
|
|
6 |
|
|
mangánové baktérie – kultivačne |
KTJ/100 ml |
|
|
0 |
|
|
živé organizmy |
i/ml |
0 |
0 |
0 |
0 |
|
bezfarebné bičíkovce |
i/ml |
10 |
50 |
0 |
0 |
|
mŕtve organizmy |
i/ml |
30 |
30 |
0 |
0 |
|
abiosestón |
% |
10 |
10 |
2 |
3 |
|
železité a mangánové baktérie |
% |
10 |
20 |
0 |
0 |
|
Ciliata |
i/ml |
|
|
|
0 |
|
nasýtenie vody kyslíkom |
% |
>50 |
>50 |
|
34 |
|
rozpustený kyslík |
mg/l |
|
|
|
3,7 |
|
kyanidy |
mg/l |
|
0,03 |
|
<0,02 |
|
striebro |
mg/l |
0,05 |
0,05 |
|
0,00012 |
|
selén |
mg/l |
0,01 |
0,01 |
|
<0,0004 |
|
vanád |
mg/l |
|
|
|
<0,001 |
|
bromdichlórmetán |
mg/l |
0,015 |
0,015 |
|
<0,015 |
|
styrén |
μg/l |
20 |
20 |
|
<0,02 |
|
2,4-dichlorófenol |
mg/l |
0,002 |
0,002 |
|
<0,00001 |
|
2,4,6-trichlórfenol |
mg/l |
0,01 |
0,01 |
|
<0,001 |
|
celková objemová alfa-aktivita |
Bq/l |
|
0,1 |
|
0 |
|
celková objemová beta-aktivita |
Bq/l |
|
1 |
|
0,05 |
|
objemová aktivita radónu 222 |
Bq/l |
|
50 |
|
50,4 |
Vegetácia lužných ekosystémov (Šomšák, 1999, 2001) je viazaná na hydropedologické podmienky vytvorené Dunajom, najmä v najmladšom období holocénu. Platí to o všetkých typoch rastlinstva, t.j. od vyslovene vodných fytocenóz, cez močiarne a brehové typy až po kriačinnú a lesnú vegetáciu. Je to veľmi dynamická vegetácia, ktorá sa v porovnaní s klimazonálnymi typmi rastlinstva dokáže prispôsobiť meniacim sa podmienkam vodného režimu v priebehu relatívne krátkeho obdobia a následne vytvoriť stabilné ekosystémy.
Napriamenie rieky a výstavba ochranných hrádzí podstatne zasiahli do pôvodného režimu vôd Dunaja a spôsobili preformovanie sa rastlinstva. Odstavenie vôd pretekajúcich okolo Malého Dunaja podnietilo zarastanie mnohých mŕtvych ramien, čo vyústilo do vzniku zaujímavých spoločenstiev hydro-hygrofytov. Na ich floristické bohatstvo, i keď už značne antropicky pozmenené, poukázal Hejný (1960). Žiaľ, mnohé z nich rozsiahlymi odvodňovacími prácami Žitného ostrova koncom päťdesiatych rokov 20. storočia zanikli.
K druhým významným zmenám došlo v medzihrádzovom priestore napriameného nového toku Dunaja a jeho ramien, v terajšom inundačnom území, Obr. III.15. Záplavy sa tu znásobili a rýchlosť prúdenia a búrlivosť ich vôd narástla. Pretrvávanie takého stavu po niekoľko desaťročí začalo selektívne pôsobiť na celé rastlinstvo, preukazne však na lužné dreviny. Tvrdé listnáče ako je dub letný (Quercus robur), menej i jaseň úzkolistý (Fraxinus angustifolia) neznášali častú prítomnosť takýchto vôd a z inundačných území medzihrádzového priestoru postupne vymizli. Na ich miesto nastúpili vŕby, a to predovšetkým vŕba biela (Salix alba), menej i vŕba krehká (Salix fragilis) a topole (Populus nigra, Populus alba, Populus x canescens), (Šomšák, 1999, 2001). Vŕbovo-topoľové lesy tu existovali aj predtým, avšak v podstatne menšej rozlohe.
Zmena rastlinstva sa dotkla aj nelesnej a vodnej vegetácie. Niektoré niekdajšie úseky mŕtvych ramien sa v inundačnom území s častejšími a mohutnejšími záplavami stali prietočnými (eupotamon) a nedovoľovali vznik a existenciu rastlinstva stojatých vôd. Vodná vegetácia mala zabezpečené existenčné podmienky len v ramenách, ktoré boli iba občas prietočné (plesiopotamal) – Oťaheľová (2001). Jej rozvoj ale rapídne začal vo vnútri nížiny a v ramenách odstavených ochrannými hrádzami (paleopotamal). Vývoj brehovej (litorálnej) vegetácie nadväzujúcej na stojaté vody tu bol plošne výraznejší, ale s mnohými prvkami sezónnosti, vyplývajúcimi z kolísania hladín v ramenách (Kubalová, 2001).
K takýmto pomerom inundácie sa lužné ekosystémy v priebehu 1950. až 1970. rokov dokonale prispôsobili. Preto prvú komplexnú informáciu venovanú pôdne ekologickým pomerom a lužným lesom Podunajskej nížiny (Jurko, 1958) možno považovať za takú, ktorá charakterizuje približne 50. roky 20. storočia, kedy ešte stav narušenia „prirodzeného“ prostredia nebol tak intenzívny. Pri posudzovaní ďalších zmien tento stav vo väčšine prípadov považujeme za východiskový, predstavujúci vegetáciu blízku prírodnej (nie však pôvodnú v pravom slova zmysle). Žiaľ o vodných a močiarnych spoločenstvách inundácií z tohto obdobia máme len strohé poznámky (Jurko, 1958).
Pokles hladín podzemných vôd po rozsiahlych úpravách koryta Dunaja, ešte pred výstavbou Sústavy vodných diel, bol citeľný zvlášť v hornej časti Žitného ostrova pod Bratislavou, najmä v biskupickom regióne, smerom k Čunovu klesal, ale citeľný bol aj v terajšom inundačnom území, Obr. III.18, Obr. III.24. Keďže v inundácii sa už dávno, v rokoch úprav koryta Dunaja, rozkladali rozsiahle monokultúry šľachtených euamerických, ale aj našich topoľov, dopad poklesu podzemných vôd na lesy a ich spoločenstvá bol menej signifikantný. Už vtedy (dávno pred výstavbou vodného diela) však došlo k poklesu prírastku lesných drevín v úzkom pobrežnom páse Dunaja, najmä na štrkových brehoch (drenážny efekt Dunaja – Šomšák et al., 1995, FNSCU, 1995). Citeľnejší dopad mala úprava koryta na vodné močiarne typy vegetácie ramennej sústavy inundačného územia. Väčšina ramien bola prietočná len pri zvýšených prietokoch vody v Dunaji. Nepravidelnosťou prietokov v ramenách však došlo k plošným zmenám medzi spoločenstvami typu plesiopotamon, parapotamon a eupotamon. Podľa osobných skúseností Šomšáka z tohto obdobia tu prevládali spoločenstvá typu plesiopotamon. Tento predpoklad potvrdzujú i novšie výskumy (Oťahelová, 2001; Kubalová, 2001; Svobodová, 1994; Matis, 2001).
Pripravovaná výstavba Sústavy vodných diel Gabčíkovo – Nagymaros si vyžiadala podrobný floristický výskum celého Podunajska. Touto inventarizáciou tu bolo zistených 959 taxónov cievnatých rastlín. Rozbor viazanosti na stanovištné (fytocenotické) skupiny ukazuje (Šomšák, 1999), že z tohto počtu len jedna tretina (311 taxónov) je takých, ktorých život limitujú podzemné a záplavové vody. Sú to vodné a močiarne rastliny (97 druhov), brehové populácie (litorálna, limózna a terestrická ekofáza) so 70 druhmi a nakoniec rastliny, ktorých životný cyklus je viazaný na lužné lesy a kriačiny (194 taxónov). Medzi ostatnými je však obrovský podiel takých druhov, ktoré dokážu a v skutočnosti aj existujú i vo fytocenózach mimo aluviálnych nív (Urtica, Glechoma, Alliaria, Symphytum, Rubus, Poa, Viola, Gagea, Sambucus, Lythrum, Lysimachia a mnohé iné).
Ostatné druhy známe zo spomínanej inventarizácie sa viažu na také stanovištia, ktoré nie sú a ani neboli ovplyvňované vodami Dunaja. Sú to napr. druhy xerotermných štrkov (180 taxónov), populácie ruderálnych stanovíšť (190 populácií), obilnín a okopanín (89 taxónov), introdukované druhy (72 taxónov) a neofytné populácie (43 druhov). Stručne povedané až 68,7 % zistených druhov tu existuje bez závislosti na vodách Dunaja (Šomšák, 1999; FNSCU, 1995). Z ekosozologického hľadiska je však najvýznamnejších práve zvyšných 31 %.
Popri analýze fytogenofondu boli navrhnuté i určité opatrenia tak, aby výstavbou vodného diela došlo len k minimálnym stratám (Bertová et al., 1986). Na prvom mieste išlo o rozšírenie počtu maloplošných chránených území, ktoré popri existujúcich prírodných rezerváciách budú chrániť celkové stanovištné podmienky vrátane tej vegetácie, ktorej súčasťou sú príslušné druhové populácie. Na úseku Bratislava – Hrušov bolo navrhnutých 9 maloplošných chránených území (Bertová et al., 1986), v terajšej inundácii 7 (Dunajská Sihoť, Šulianské ramená, Kráľovská lúka, Bodícke ramená, Bačianske ramená pri Bake, Istragov a Riečina (Erčéd)). Pre sledovanie osudu flóry po výstavbe VD v úseku inundácie boli vyzdvihnuté dva druhy, a to bleduľa letná (Leucojum aestivum) a kotvica kužeľoplodá (Trapa conocarpa). Posledný predstavuje nový druh pre flóru Slovenska a jeho výskyt bol zaznamenaný v mŕtvom ramene pri Kráľovskej lúke /Bodíky).
Napriek týmto opatreniam v priebehu prípravných prác pred výstavbou VD došlo k citeľnému zásahu do genofondu rastlín, a to na prvom mieste k zmenšeniu areálu mnohých populácií zabratím pôdneho (lesného i nelesného) fondu pre zdrž, derivačný a odtokový kanál (pôdny fond), priesakové kanály a i.
Údaje o flóre tohoto územia sa tesne pred plánovaným uvedením vodného diela Gabčíkovo do prevádzky spresnili podrobnou inventarizáciou spojenou so zakladaním monitorovacích plôch. Uherčíková (2001) uvádza 760 taxónov. Ako uvádza autorka, už pred spustením prevádzky VD Gabčíkovo tu chýbali viaceré taxóny vzácnych a ohrozených rastlín, ktoré tu boli zaznamenané v 50. rokoch 20. storočia (Hottonia palustris, Gratiola officinalis, Senecio paludosus, Veronica catenata, Sagittaria sagittifolia). Zachovali sa však lokality s početným výskytom úplne chráneného druhu Leucojum aestivum.
Lesné hospodárstvo realizuje svoje zámery v inundačnej oblasti na rozlohe okolo 3100 ha lesa. Táto rozloha bola od 1960-tych rokov len nepatrne zväčšená. Od 1960. rokov dochádzalo k zakladaniu veľkoplošných monokultúr do vopred pripravenej pôdy. V mnohých prípadoch sa zalesnili aj bývalé mŕtve ramená, do ktorých sa počas vytláčania pňov a iných pozostatkov po ťažbe dreva nahrnula skrývka. K hlavným drevinám, ako je vŕba biela, vŕba krehká, topoľ čierny, topoľ biely, topoľ sivý sa už v 60. rokoch pridávali kultúry cudzokrajných topoľov. Už okolo roku 1956 sa ich rozloha pohybovala okolo 27 % z existujúcej rozlohy lesov (Jurko, 1958). Od roku 1956 sa ich plošný podiel prudko zvyšoval a už okolo roku 1981 dosahovala v dunajských lužných lesoch okolo 80 % (Vojtuš, 1986). V prvých začiatkoch to boli kultivary Populus deltoides – „Monilifera“ a Populus x euroamericana – „Robusta“ a neskôr i rajonizovaný klon „I-214“ vyšľachtený v Taliansku (Neštický, Varga, 2001).
Lesnícky výskumný ústav Zvolen – Výskumná stanica Gabčíkovo už od roku 1956 zakladal v tomto území pokusné plochy. Zo získaných poznatkov rozmanitých klonov topoľov a vŕb od začiatkov ich cieleného pestovania až do prehradenia Dunaja sa potvrdil predpoklad, že ide o dreviny a porasty tolerantné voči menším zmenám vodného režimu (Varga, 1993; Neštický, Varga, 2001). Za pestovanie euroamerických topoľov a ich kultivarov sa prihovárala aj nezvyčajne vysoká produkcia drevnej hmoty. Prvé merania signalizovali ročný prírastok až 25 m3/ha. Pokles prírastku drevnej hmoty signalizovali len porasty ľavobrežného pásu Dunaja medzi Čunovom a Sapom. Dendroekologické bádania tohto úseku však potvrdili pokles prírastkov dávno pred prehradením Dunaja (bagrovanie dna rieky – Šomšák et al., 1995).
Na základe dostupných literárnych prameňov, ale aj z autentických poznatkov Šomšáka z týchto rokov, možno stav inundačného územia Dobrohošť – Sap pred prehradením Dunaja charakterizovať nasledovne:
Rozsiahly komplex vŕbovo-topoľových lesov (Salici-Populetum) so všetkými subtypmi prispôsobenými rôznym hladinám podzemných vôd, avšak s podstatne zmenenou drevinovou skladbou, kde pôvodné dreviny boli nahradené veľkoplošnými monokultúrami euroamerických topoľov iných šľachtencov.
Zvyšky pôvodných (prírodných) vŕbovo-topoľových spoločenstiev zachovaných v terénnych depresiách a na ťažko dostupných ostrovčekoch (asi 10 % porastovej plochy).
Nepatrná (mozaikovitá) rozloha spoločenstiev prechodného lužného lesa (Fraxino angustifoliae – Populetum albae) v podiele asi 11,4 % porastovej plochy.
Úzky pobrežný pás degradujúcich lesných porastov na štrkovitých laviciach fungujúcich ako drenážny systém, vyvolaný prehĺbením koryta po ťažbe štrku z Dunaja (asi 3 % porastovej plochy).
Vysokostebelnatá močiarna vegetácia na čiastočne zazemnených, len občas prietočných ramenách.
Nepatrná rozloha vodno-močiarnej vegetácie typu paleopotamon na odrezaných ramenách (Istragov, Erčéd, Kráľovská lúka).
Sezónna, jednoročná vegetácia obnažených brehov ramien závislá na dobe obmývania.
Terestrické živočíšne spoločenstvá inundačnej oblasti (ale aj ďalších častí Podunajska, snáď s výnimkou xerotermných) sa v minulosti monitorovali len sporadicky. Väčšina prác z obdobia pred zahájením monitorovania v roku 1990 je orientovaná faunisticky, prípadne len na základné ekologické otázky, napríklad väzbu na pôvodné, neraz už pozmenené alebo zaniknuté biotopy (Štepanovičová, 1995). Pri definovaní pôvodného stavu spoločenstiev terestrických živočíchov Podunajska možno vychádzať len čiastočne z údajov získaných priamo z tejto oblasti. Aj vtedajší stav už bol výsledkom vyše storočného radikálneho zasahovania do vodného režimu Dunaja a jeho okolia. Konzistentné dáta boli získané až od začiatku monitorovania bioty v roku 1990, v súvislosti s výstavbou a prevádzkou vodného diela Gabčíkovo (Jedlička et al., 1999). Za relatívne pôvodný stav môžeme považovať zoocenózy zvyškov lužných lesov v medzihrádzovom priestore v úseku Sap – Čičov, ktoré bývajú aj v súčasnosti zaplavované a zásobované priesakovou vodou, a štruktúrou sa najviac podobajú pôvodným spoločenstvám najvlhších typov mäkkých lužných lesov s prevahou polyhygrofilných druhov adaptovaných na podmienky záplav.
Terestrická fauna a zoocenózy v inundácii (Jedlička, Országh, Čejka, Darolová, Kulfan, Mikulíček, Šustek, Žiak, 1999; OPTIMALIZÁCIA, 2003) sú typické pre inundačné územie Dunaja medzi protipovodňovými hrádzami, bohatou sieťou riečnych ramien a stojatých vôd, súvisiacou s eróziou a sedimentáciou nánosov, meandrovaním rieky a častými záplavami Táto dynamika je špecifická pre komplex ekosystémov vodného, pôdneho a suchozemského prostredia a tomu zodpovedajúcich ekotonov v terestrickej časti, s výskytom nasledujúcich zväzov a asociácií: Phragmition, Magnocaricion elatae, Caricion gracilis, Oenanthion aquaticae, Elatino-Eleocharition ovatae, Chelidonio-Robinion, Lolio-Potentillion, Salicion albae, Ulmenion, Asparago-Crataegetum. Jednotlivé spoločenstvá predstavujú katenu (zákonitý sled typov pôd a ekosystémov, v tomto prípade viazaný na typickú zmenu hĺbky hladiny podzemnej vody a geologické zloženie zóny medzi terénom a hladinou podzemnej vody – kapilárne vzlínanie) pozdĺž vlhkostného gradientu súvisiaceho s hĺbkou a kolísaním hladiny podzemných vôd a existenciou sezónnych záplav.
V plochých nížinných alúviách sa vytvárali charakteristické spoločenstvá s vysokými nárokmi na vlhkosť, ktoré vďaka šírke nivy neboli, v relatívne širokom pásme, infiltrované mezohygrofilnými druhmi. K vzájomnému prelínaniu druhov dochádzalo až vo väčšej vzdialenosti od toku, prípadne až na samotnom okraji nivy. Šírku tohto prechodného pásma určovala konfigurácia terénu a vzájomný kompetičný tlak druhov rôznych ekologických skupín. Poloha tohto pásma sa dynamicky menila podľa výkyvov vodnatosti a meandrovania toku. Táto situácia umožňovala, podľa zmien polohy koryta, rozsahu záplav a výšky hladiny podzemnej vody na jednom mieste rýchle obmieňanie rôznych, ale vždy prírodných, spoločenstiev. Takáto situácia bola v dávnej minulosti typická pre rozsiahle nížinné územia a aj pre Podunajskú nížinu.
Formovanie prirodzeného rozšírenia spoločenstiev v závislosti od vlhkostného gradientu územia obmedzujú protipovodňové hrádze a poľnohospodárska činnosť za hrádzami. Tieto odrezali za hrádzami časť lesných porastov od prirodzeného záplavového režimu. Preto sukcesia spoločenstiev v mimohrádzovom priestore má predpoklady smerovať k mezohygrofilným spoločenstvám lesných geobiocenóz normálneho hygrického radu, prípadne pri narušení integrity porastu k spoločenstvám nelesných ekosystémov. Naopak časť spoločenstiev v užšom medzihrádzovom priestore je vystavená intenzívnejšiemu pôsobeniu záplav usmernených do pomerne úzkeho koridoru a zbavených možnosti rozliať sa do odľahlejších časti alúvia. Jeden i druhý stav sa teda odlišuje aspoň na časti sledovaného územia od stavu, ktorý tu bol v prírodnej, alebo človekom len zanedbateľne zmenenej krajine. Pre pôvodnú krajinu, hlavne kvôli menším záplavovým výškam, bol charakteristický aj výskyt tvrdých lesných listnáčov ako je dub letný (Quercus robur), menej i jaseň úzkolistý (Fraxinus angustifolia). Na ich miesto nastúpili vŕby a to predovšetkým vŕba biela (Salix alba), menej i vŕba krehká (Salix fragilis) a topole (Populus nigra, Populus alba, Populus x canescens). Vŕbovo-topoľové lesy tu boli aj predtým, avšak v podstatne menšej rozlohe.
Zoocenózy ako spoločenstvá konzumentov a producentov sú v celom sledovanom území viazané na (Jedlička et al., 1999):
a) amfibické a prechodné spoločenstvá asociácií Rorippo - Agrostietum stoloniferae, Rorippo amphibiae - Oenanthetum aquaticeae, Eleocharitetum palustris, Glycerietum maximae, Phalaridetum arundinaceae, Phragmitetum communis a Potametum perfoliati, Caricetum gracilis,
b) mäkké lužné lesy Salici-Populetum v rôznych podtypoch a stupni pôvodnosti,
c) prechodné lužné lesy Fraxino angustifoliae – Populetum albae,
d) tvrdé lužné lesy Fraxino angustifoliae – Ulmetum (platí len pre časť lesov nad prehradením Dunaja),
e) dunajskú lesostep Asparago-Crataegetum.
Malakocenózy sú taxocenózy, ktoré veľmi dobre dokumentujú nielen stav ale aj vývoj zoocenóz. Je známe, že taxocenózy mäkkýšov (Mollusca) prešli v kvartéri zložitým sekulárnym vývojom (Ložek, 1955). Súčasné spoločenstvá sa formovali postupne v mladšom holocéne (Cheben et al., 2001), pravdepodobne na omnoho rozsiahlejšom území, takže dnešný pás v medzihrádzovom priestore je len ich zvyškom. Podľa doterajších nálezov holocénnej malakofauny je pravdepodobné, že približne v oblasti dnešnej obce Vrakúň kedysi prebiehala hranica oddeľujúca široký pás zaplavovaných luhov od vyvýšeného, suchšieho, čiastočne stepného vnútra Žitného ostrova. Inundačné územie na dnešnej slovenskej strane malo teda v mladšom holocéne šírku približne 10 km (dnes sú to v najširšom mieste pri Bake 3 km) (Ložek, 1955, 1964). Analýzy tanatocenóz sedimentov Dunaja vo Vlčom Hrdle pri Bratislave kombinované s absolútnym datovaním veku sedimentov (Pišút, 2000; Čejka, 2000) umožnili zrekonštruovať kompletnú sukcesnú líniu lužného lesa od iniciálneho štádia lužného lesa až po dnešný prechodný lužný les (obdobie r. 1791-1999) a potvrdili výskyt podobných typov lužného lesa, aké sa na Podunajsku vyskytujú v súčasnosti, a tanatocenózy, ktoré štruktúrou zodpovedajú približne dnešným zvyškom najzachovalejších lesných malakocenóz v oblasti medzi Dobrohošťou a Sapom. Celková plocha najvlhších typov mäkkých lužných lesov s príslušnými malakocenózami bola v minulosti oveľa väčšia ako dnes, s prirodzenejšou štruktúrou porastov, čo je zreteľne vidieť dokonca už pri porovnaní fytocenologických zápisov zo 1940-1950-tych rokov so súčasnými (Uherčíková, 1995, 1998).
Charakteristické, hlavne pre iniciálne štádiá tzv. mäkkého luhu a iné stanovištia s vysokou pôdnou vlhkosťou, sú najmä výrazne vlhkomilné druhy ulitníkov Succinea putris, Oxyloma elegans, Zonitoides nitidus a Pseudotrichia rubiginosa. Diferenciačnými druhmi vlhkých typov mäkkého lužného lesa (asoc. Salici-Populetum myosotidetosum až Salici-Populetum typicum Jurko, 1958) sú, okrem vyššie uvedených druhov aj polyhygrofilný Carychium minimum a lesné hygrofilné druhy Arianta arbustorum, Vitrea crystallina a sčasti aj Urticicola umbrosus. Pre tzv. prechodný až tvrdý luh (as. Fraxino-Populetum, Fraxino-Ulmetum) je zase typická dominancia prevažne lesných mezohygrofilných druhov, ktoré neznášajú ničivý vplyv záplav a dlhodobo podmáčanú pôdu (Aegopinella nitens, Cochlodina laminata, Semilimax semilimax, Alinda biplicata, Monachoides incarnatus, Petasina unidentata, Clausilia pumila, čiastočne aj Carychium tridentatum). V taxocenózach sú zastúpené aj skupiny druhov, ktoré sú viazané na vyslovene nelesné stanovištia, alebo riedko zapojené porasty stromov či krov (Vallonia pulchella, V. costata, Euomphalia strigella, Cepaea vindobonensis a Xerolenta obvia).
Analogicky je možné predpokladať podobné pomery (pokiaľ ide o zastúpenie jednotlivých ekoelementov najmä vo vzťahu k vlhkosti, vode, zatieneniu atď.) aj v iných taxocenózach terestrických živočíchov.
Vo faune suchozemských rovnakonožcov (Oniscidea) z Podunajska v dosahu Vodného diela Gabčíkovo bolo z obdobia 1986-1990 zistených 16 druhov (Flasarová, 1999), najpočetnejším bol eurytopný Trachelipus rathkei.
Stonožkám (Chilopoda) Podunajska bola v období pred zahájením výstavby VD Gabčíkovo venovaná len okrajová pozornosť a podobne ako u predchádzajúcej skupiny netýkajú sa priamo územia tejto štúdie, ale jeho okolia (Gulička, 1957; Čarnogurský et al., 1994).
Za východiskový stav taxocenóz stonožiek sa môže do istej miery považovať druhové zloženie zistené v úseku Dobrohošť – Sap pred rokom 1993. Na monitorovacích plochách bolo zaznamenaných 9 – 14 druhov. Získané údaje o spoločenstvách suchozemských rovnakonožcov (Oniscidea) a stonožiek (Chilopoda) sú veľmi blízke až totožné s údajmi z lesov alúvií Moravy a Dyje na moravskej i rakúskej strane (Tajovský, 1999; Tuf, 2000; Zulka, 1999), môžeme ich považovať za dostatočne charakteristické.
V Dobrohošti - Dunajských krivinách z 13 zistených druhov k eudominantom patril eurytopný Lithobius forficatus, z okolia sem prenikal Lithobius mutabilis. Z pravidelne sa vyskytujúcich druhov, ktoré sú indikátormi vyššieho stupňa pôdnej vlhkosti (od 20 % - 30 %) boli zistené mezohygrofilné Lithobius curtipes, L. crassipes, Geophilus flavus. Lithobius microps tu už po roku 1991 zistený nebol.
Spoločenstvo stonožiek Bodíckej brány tvorilo 13 druhov, medzi ktoré patrili mezohygrofilné L. crassipes a L. curtipes s vyššími hodnotami dominancie (17 % - 18 %) a hygrofilný L. agilis, náročný na pomerne vysoký stupeň pôdnej vlhkosti. Tretinu druhov však predstavovali eurytopné druhy L. forficatus, L. erythrocephalus a L. mutabilis, ktoré boli a sú zastúpené po celom sledovanom území Podunajska.
Taxocenózu stonožiek Kráľovskej lúky tvorilo pred prehradením 14 druhov, ku ktorým patrili typický ripikolný a hygrofilný Lamyctes emarginatus a hygrofilné druhy L. agilis a L. microps. Eudominantným druhom bol Lamyctes emarginatus, dominantnými Lithobius aeruginosus a L. curtipes. Spoločenstvo stonožiek Kráľovskej lúky pred prehradením Dunaja zaraďujeme medzi najbohatšie. Za obdobie rokov 1991 – 1997 tu bolo zistených 17 druhov. V prvom roku po prehradení (1993) počet zaznamenaných druhov bol zreteľne nižší oproti rokom predchádzajúcim. Absentujúce druhy sa opäť objavili až v roku 1997, naviac v tomto roku bol po prvýkrát zistený Lithobius pelidnus, druh s centrom rozšírenia vo východnej Európe preferujúci zalesnené územia.
V spoločenstve stonožiek Istragova bolo zastúpených 9 druhov, z ktorých medzi eudominantné patrili eurytopné Lithobius forficatus a L. mutabilis. Z typických mezohygrofilných druhov boli vysokým percentom zastúpené tiež L. aeruginosus, L. crassipes, L. curtipes a hypogeicky žijúci druh Pachymerium ferrugineum. Pred prehradením Dunaja nechýbal ani hygrofilný druh L. agilis zistený v rokoch 1991 a 1992.
Z územia, na ktorom je v súčasnosti vybudované VD Gabčíkovo a jeho najbližšieho okolia viacerí autori (Brtek, Rotschein, 1964; Šiška 1983; Štepanovičová, Lapková, 1984; Bulánková, 1995; Štepanovičová, 1989, 1991) zistili pred začatím výstavby výskyt 177 druhov bzdôch (Heteroptera).
V epigeickej taxocenóze bystruškovitých (Coleoptera: Carabidae) v podunajských lužných lesoch s pravidelnými záplavami prevládali s vysokou dominanciou vlhkomilné druhy. Na Dunajských krivinách to boli hlavne Agonum moestum, Oxypselaphus obscurus, Patrobus atrorufus a Platynus assimilis. Jednotlivo sa objavovali aj ďalšie vlhkomilné alebo ripikolné druhy (napr. Bembidion dentellum, Bembidion biguttatum, Oodes helopioides). Podobne v Bodíckej bráne výrazne prevažovali vlhkomilné druhy P. assimilis, P. atrorufus, O. obscurus sprevádzané ďalšími početne zastúpenými vlhkostne menej náročnejšími, resp. tolerantnejšími druhmi Asaphidion flavipes, Carabus granulatus, Pterostichus strenuus, Clivina fossor. Významná bola aj prítomnosť vlhkomilných druhov B. dentellum a Badister sodalis.
Na Kráľovskej lúke, kde bolo spoločenstvo sledované už od roku 1987, prevažovali paludikolné A. moestum, vlhkomilné druhy P. atrorufus, P. assimilis a relatívne tolerantné druhy P. strenuus , C. granulatus a O. obscurus. Vysokej vlhkosti, resp. podmáčaniu zodpovedala prítomnosť Europhilus fuliginosus, Europhilus micans, Oodes helopioides. V roku 1989 sa abundancia všetkých vlhkomilných druhov okrem A. moestum zvýšila. Veľmi prudko vzrástlo zastúpenie A. flavipes (z 2 na 248 jedincov). V rokoch 1990-1991 bol celkový úlovok v dôsledku zaplavenia lokality nižší (z 1348 jedincov v r. 1989 klesol na 727, resp. 372 jedincov), ale štruktúra spoločenstva sa výrazne priblížila pôvodnému stavu z roku 1987. Zaplavenie postihlo aj dva vlhkomilné, ale nie vyslovene paludikolné druhy P. atrorufus a P. assimile.
Na Istragove v roku 1989 výrazne prevažovali dva drobné vlhkomilné druhy bystruškovitých A. flavipes a Bembidion femoratum, výrazné zastúpenie mali aj ďalšie vlhkomilné druhy P. assimile, P. strenuus, C. granulatus, P. atrorufus. Ekologicky významná bola prítomnosť druhov Europhilus fuliginosus a E. micans. Vysoké zastúpenie vlhkostne menej náročných druhov ešte pred podstatnými zmenami hydrologického režimu odráža prevahu silne piesčitej pôdy na veľkej časti sledovanej plochy.
Načrtnutý stav naznačuje, že taxocenózy bystruškovitých predstavovali charakteristické spoločenstvá druhov s vysokými nárokmi na vlhkosť, ktoré neboli v podstatnej miere infiltrované mezohygrofilnými druhmi obývajúcimi geobiocenózy normálneho hydrického radu. Neboli ani výrazne ovplyvnené druhovou skladbou porastov, ktorá bola v ostatných desaťročiach silne narušená náhradou pôvodných porastov topoľovými kultúrami. Príčinou je skutočnosť, že pre hygrofilné druhy lužných lesov je rozhodujúca prítomnosť v podstate akéhokoľvek hustého vegetačného krytu, ktorý zabraňuje vysušovaniu a prehrievaniu hrabanky priamym slnečným žiarením. Takéto podmienky pre taxocenózy bystruškovitých boli schopné vytvárať tak topoľové kultúry staršie ako 10 rokov, t. j. počas prevažnej časti svojej existencie, ako aj vysoký bylinný porast, ktorý sa (bez ohľadu na svoje floristicky nevyhovujúce druhové zloženie) vyvinul na rúbaniskách v podmienkach dostatočného zásobenia pôdy vlhkosťou.
Pre semiakvatické, amfibické a prechodné živočíšne taxocenózy je pomerne charakteristickým javom ich väzba nielen na vegetáciu ako potravnú bázu, ale aj viazanosť na vodný režim; jeho nepravidelné zmeny s následnou sukcesiou sa prejavujú na nestabilite zloženia taxocenóz a ich veľkých medziročných zmenách. To dokumentuje situácia taxocenóz fytofágnych Curculionidae (Coleoptera) brehových vegetačných formácií v systéme dunajských ramien a hlavného toku Dunaja. Staršie údaje sú v širšie koncipovaných štúdiách (Majzlan, Rychlík, 1982; Majzlan, 1988, 1990; Kodada, Majzlan, 1991) a neskôr boli aj monitorované. Z uvedených prác vyplýva, že pobrežné územia niektorých skúmaných ramien boli už pred prehradením Dunaja do značnej miery aridizované. Odrazilo sa to aj na pomernom zastúpení eurytopných a stenotopných druhov: hygrofilných a paludikolných na jednej a druhov xerofilných a na biotop nenáročných na strane druhej. Je možné sa oprávnene domnievať, že spoločenstvo nosáčikov (Curculionidea) zistené v rokoch po prehradení Dunaja (1992) žilo na skúmanom území v pobrežnej vegetácii ramien vnútrozemskej delty aj pred jeho prehradením. Z vyschnutých ramien sa táto taxocenóza stiahla do menších enkláv, v ktorých prežívala. S časom prehradenia Dunaja súvisí šírenie smerom na sever hygrofilného a ripikolného nosáčika Bagous bagdatensis. Jeho lokality na území Slovenska predstavujú dosiaľ známu severnú hranicu rozšírenia. Ripikolný a akvikolný Dicranthus majzlani, na území Slovenska aj v celej Európe kriticky ohrozený, je indikátorom prírodne zachovalých stojatých a polotečúcich nížinných vôd a vyžaduje vyššiu hladinu vody v ramennom systéme. Obidva druhy neboli do roku 1992 z tohto územia známe.
V priebehu monitorovania sa zistilo, že zatiaľ čo v komplexe pobrežných rastlinných spoločenstiev (asociácie Rorippo-Agrostietum stoloniferae, Phalaridetum arundinaceae, Rorippo amphibiae-Oenanthetum aquaticae, Glycerietum maximae, Phragmitetum communis, Caricetum gracilis) bolo v taxocenóze Curculionidae zistených 49 druhov, z toho 13 každoročne (Sitona macularis, Sitona suturalis, Bagous collignensis, B. glabrirostris, Tanysphyrus lemnae, Rhinoncus albicinctus, R. perpendicularis, R. inconspectus, Poophagus sisymbrii, Tapinotus sellatus, Nanophyes brevis, N. globiformis, N. marmoratus) s vyrovnaným pomerom hygrofilných druhov viazaných na pobrežnú vegetáciu, ako aj druhov viazaných na rastliny vodnej hladiny so signifikantnou prevahou charakteristických, stenotopných a hygrofilných druhov, tak vo vysychajúcom slepom ramene Dunaja v lužnom lese (Salici-Populetum) s asociáciou Phragmitetum communis s väčším množstvom vody iba v jarnom období (apríl a máj) bolo v taxocenóze Curculionidae zistených 39 druhov, pričom ani jeden druh sa nevyskytoval každoročne. Eudominantným bol sprievodný druh Nedyus quadrimaculatus.
Z územia boli dávnejšie pomerne dobre známe taxocenózy suchozemských resp. amfibických stavovcov. Na území bol známy výskyt 12 taxónov obojživelníkov, z nich Triturus dobrogicus a Rana ridibunda sú v kategórii ohrozených (EN), Triturus vulgaris a Rana lessonae v kategórii zraniteľných (VU), všetky ostatné v kategórii rizikových (LR) druhov. Z 12 druhov plazov známych z územia Slovenska sa tu vyskytuje 9, z toho 7 chránených, 3 v kategórii zraniteľných (VU: Coronella austriaca, Natrix tesselata, Lacerta viridis), ostatné v kategórii rizikových (LR) druhov.
Z ornitologického hľadiska predstavovali podunajské lužné lesy spolu s ramenným systémom Dunaja v minulosti územie s vysokou diverzitou a denzitou druhov, kde hniezdili viaceré vzácne a ohrozené druhy vtákov (Balát, 1963; Rybanič, 1999). Hniezdnu ornitocenózu podunajských lužných lesov v 1970-tych a 1980-tych rokoch tvorilo 103 druhov vtákov. Z významných hniezdičov to boli predovšetkým haja tmavá (Milvus migrans - VU) a chochlačka bielooká (Aythya nyroca - EN), ktoré tu vytvárali hniezdne populácie celoslovenského významu, ďalej bučiačik močiarny (Ixobrychus minutus VU), bocian čierny (Ciconia nigra), včelár lesný (Pernis apivorus), rybárik riečny (Alcedo atthis), ďateľ prostredný (Dendrocopos medius), ktoré tu vytvárali hniezdne populácie nadregionálneho významu. Z celkového počtu 103 hniezdičov boli 3 druhy ohrozené (EN: Ardea purpurea, Aythya nyroca, Coracias garrulus), a 4 zraniteľné (VU: Ixobrychus minutus, Milvus migrans, Nycticorax nycticorax, Upupa epops).
Okrem lužných lesov dôležitých pre hniezdiče je Dunaj významnou trasou migrácie vodného vtáctva. Na hlavnom toku Dunaja zimovalo v jednotlivých rokoch 25-30 druhov vtákov (Kalivodová, Darolová, 1998, Áč et al., 1996). Medzi dominantných hibernantov patrili Anas platyrhynchos a Bucephala clangula.
Vo faune cicavcov (Mammalia) bolo zistených 49 druhov. Bobor (Castor fiber), na území vyhynutý, sa v súčasnosti po reštitúcii v Rakúsku spontánne šíri, nenachádza však v študovanom území optimálne podmienky. V taxocenóze drobných zemných cicavcov, tvorenej druhmi Sorex araneus, Apodemus flavicollis, Clethrionomys glareolus, Sorex minutus, Crocidura leucodon, Crocidura suaveolens, Microtus arvalis, Microtus oeconomus, Pitymys subterraneus, Apodemus sylvaticus, Micromys minutus dochádza v závislosti najmä na vlhkostnom gradiente k zmene štruktúry dominancie. V mäkkom lužnom lese sú eudominantné Sorex araneus, Apodemus flavicollis a Clethrionomys glareolus, s posunom na vlhkostnom gradiente smerom ku xerickým podmienkam sa ich dominancia znižuje a vo zvýšenej miere sa v spoločenstve uplatňujú iné druhy vrátane tu nepôvodných Microtus arvalis a Mus musculus.
Štruktúra, dynamika a produkcia planktónu, bentosu, nektónu i neustónu verne odráža pôvodnosť tokov a kvalitu vôd. Spoločenstvá týchto organizmov charakterizujú tok a jeho ramennú sústavu. Sú významnými indikátormi zmien, schopnými reagovať na zmeny v rôznych časových rovinách, a tak sa z ich reakcií môžu vyvodzovať závery o zmenách, ktoré prebehli a aj prebiehajú. Umožňujú v základných rysoch predpokladať aj ďalší vývoj.
Technické zásahy v minulosti, s cieľom znížiť riziká povodní a zlepšiť plavebné podmienky, sa sústredili predovšetkým na odstraňovanie brodov, prehlbovanie a upravovanie dna Dunaja, regulácie a opevnenie brehov, uzatváranie ramien (Dub, Szolgay, 1966). Výsledkom toho bola výrazne zvýšená rýchlosť prúdenia v rieke a silnejšia erózia dna, výrazné zníženie a prerušenie prúdenia v pôvodne prietočných ramenách typu eupotamál a parapotamál. Pôvodná heterogenita mezohabitatov bola potlačená. Výrazne sa sťažili podmienky migrácie lariev, ale aj reprodukcie imág vodného hmyzu a iných vodných organizmov (Krno, 1995). Uzatváraním vtokov ramien a v dôsledku poklesu riečiska Dunaja sa vytvorili podmienky pre rozšírenie ramien typu plesiopotamál charakterizovaných rozvojom makrofýt, eutrofizáciou a ich postupnú premenu na vysýchajúce typy periodických mokradí. Vybudovanie protipovodňových hrádzí spôsobilo stratu spojenia ramien v mimohrádzovom priestore s hlavným tokom a ich premenu na ramená typu paleopotamál. Takéto podmienky znamenali ústup reofilných, potamofilných a stenoekných foriem makrozoobentosu a ich nahradenie skôr eurytopnými formami.
Pre Dunaj a jeho inundáciu boli v minulosti typické viac-menej pravidelne sa opakujúce vyššie prietoky začiatkom jari v marci a začiatkom leta v júni a júli (Mucha et al., 2001). To svedčí o špecifickosti prietokového režimu tejto rieky, a preto ju nemôžeme označiť ako typicky alpský typ (Holčík et al., 1981). Výrazné jarné maximá sú výsledkom topenia snehov v predpolí Álp a Hercynika, takže ide o zmiešaný typ vysokohorského a podhorského prietokového režimu. Takémuto vodnému režimu boli podriadené aj vývinové cykly temporálnej epifauny Dunaja a jeho inundácie. Záplavy, periodické i neperiodické, boli hlavným prírodným faktorom genézy ekosystémov Dunaja (Rovný et al., 1996). Záplavy preplachovali mediál ramien, zabraňovali kolmatácii dna, najmä jarné bohato zásobovali ramená živinami, priaznivo ovplyvňovali kyslíkový i teplotný režim, umožňovali tak úspešný ontogenetický vývin hydrobiontov a v neposlednom rade uľahčovali ich pasívnu i aktívnu migráciu. Protipovodňovými hrádzami a smernými stavbami v toku bol tento režim zmenený, nastalo rýchlejšie a väčšie kolísanie hladín, zrýchlenie rýchlosti toku najmä v prehlbovanom úseku napriameného hlavného koryta, nastal pokles hladín povrchových i podzemných vôd. Od 60. rokov 20. storočia bol tento proces sprevádzaný aj postupným uzatváraním ramien, fragmentáciou vnútrodunajskej delty, poklesom frekvencie preplachovania ramien až ich úplným vyschnutím.
Faunistické údaje mikro-meiozoobentosu tejto oblasti boli publikované vo viacerých prácach (Ertl, 1966, 1970). Najviac údajov bolo známych o perifytóne, ktorý tvorili predovšetkým malé meňavky (Amoebina), niektoré druhy nálevníkov a sesilných Rotifera (Matis, Tirjaková (1992, 1995a, 1995b); Tirjaková (1992, 1999); Baláži, Tirjaková (1999); Baláži, Matis (2002)).
Makrozoobentos Dunaja, jeho ramien a periodických vôd bol predmetom štúdia viacerých autorov: Balthasara (1936, 1938); Hraběho (1941); Trpiša (1957, 1962); Raušera (1957); Lichardovej (1958); Ertlovej (1963, 1968, 1970, 1973, 1987); Brteka a Rothscheina (1964); Russev (1979); Pomichala (1984); Landu (1969); Nagya a Šporku (1990); Krna (1990); Majzlana (1992); Davida (1994); Bulánkovej (1995, 1999, 2001); Bulánkovej a Halgoša (1995, 1997); Halgoša (1995); Šíbla et al. (2001); Šporku, Krna (1995); Košela (1995a); Šporku (1980a, 1980b, 1982, 1983, 1984, 1994, 1998 ); Šporku, Nagya (1998). Z týchto prác vyplýva, že Dunaj a priľahlé inundačné územie je už vyše storočia pod silným antropickým tlakom, ktorého výsledkom je napríklad vyhynutie viac než 90 % druhov dunajských pošvatiek, viac než 50 % podeniek a viac než 30 % potočníkov (Krno a kol., 1999).
Územie Žitného ostrova bolo od dávna známe kvantitatívnym i kvalitatívnym bohatstvom komárov (neustón), ako i každoročnými kalamitami súvisiacimi s premnožením niektorých druhov. Kalamity spôsobovalo vyliatie vôd z Dunaja do inundačného územia. Trpiš (1962) udáva z územia Žitného ostrova 28 druhov komárov. Od tohto obdobia prešlo územie veľkými zmenami súvisiacimi s výraznými antropickými zásahmi (Halgoš, 1981; Halgoš, 1984).
Hlavný tok Dunaja
V rokoch 1971-1972 (Vranovský, 1974) a v rokoch 1991-1992 (Illyová, 1995; Vranovský, Illyová, 1999) dominovali medzi planktonickými vírnikmi hlavného toku spravidla euplanktonické druhy, najmä z rodov Keratella (K. cochlearis), Polyarthra (P. vulgaris, P. remata), Synchaeta (S. oblonga, S. tremula, S. stylata), Brachionus (B. calycilforus, B. angularis), a v zimnom období aj Rotaria rotatoria. V krustáceoplanktóne podobne prevládali pravé planktonty, avšak v hornej časti monitorovaného úseku (na profile Dunajské kriviny) značný podiel z celkovej abundancie pripadal aj na tychoplanktonické (litorálne a bentické) druhy, t.j. nepravé planktonty. Z perloočiek (Cladocera) dominovala Bosmina longirostris, prípadne spolu s druhom Daphnia longispina a/alebo D. cucullata, z veslonôžok (Copepoda) najčastejšie Acanthocyclops robustus, a to buď so vznášavkou Eudiaptomus gracilis alebo s druhmi Cyclops vicinus a Thermocyclops oithonoides, prípadne aj s druhom Eurytemora velox (imigrant, zistený v našom úseku Dunaja prvýkrát v roku 1991). Z tychoplanktonických perloočiek k najhojnejším patrili Alona quadrangularis a A. affinis, z veslonôžok Eucyclops serrulatus .
Ramená typu parapotamál
Pred rokom 1992 vo vodách tohto typu sa v teplej časti roka v obdobiach stagnácie prietočnosti spravidla vyvinul kvantitatívne bohatý zooplanktón tvorený výlučne euplanktonickými druhmi (Vranovský, 1974, 1985; Vranovský, Illyová, 1999). Z vírnikov dominovali viaceré druhy rodov Brachionus, Keratella, Polyarthra a Synchaeta. Z perloočiek to boli Bosmina longirostris spolu s Daphnia longispina a D. cucullata (v ramenách nad Gabčíkovom), resp. s D. cucullata, Diaphanosoma brachyurum a Moina brachiata (v Istragovskom ramene - pod Gabčíkovom). Podobne, z druhej významnej komponenty krustáceoplanktónu - veslonôžok - v stredovej zóne boli zastúpené iba pravé planktonty, najmä Thermocyclops oithonoides a Th. crassus (v Istragovskom ramene) spolu s niektorými ďalšími druhmi.
Ramená typu plesiopotamál
V hlbšom ramene na Kráľovskej lúke pri Trstenej, ale aj v plytkom zvyšku ramena na Spornej Sihoti pri Kľúčovci, v krustáceoplanktóne dominovali typické planktonty, ba v prvom roku boli jeho jedinou zložkou (Vranovský, Illyová, 1999). V ramene na Kráľovskej lúke to bola z perloočiek Diaphanosoma brachyurum alebo Bosmina longirostris, z veslonôžok Thermocyclops oithonoides, v ramene na Spornej Ssihoti medzi najčastejšie a aj v ročných priemeroch dominujúce druhy patrila perloočka Bosmina longirostris a veslonôžky Cyclops vicinus, Thermocyclops oithonoides, Th. crassus, Eudiaptomus gracilis., Eurytemora velox a tiež Acanthocyclops robustus.
Po zátopách inundačného územia sa v tomto type ramien vytváralo spoločenstvo s dominanciou euplanktonických druhov, najmä veslonôžok Cyclops vicinus a Thermocyclops crassus. V iných obdobiach tu nadobúdali kvantitatívnu prevahu druhy charakteristické pre litorál, resp. plytké občasné (temporárne) stojaté vody: z perloočiek najmä Chydorus sphaericus a Ceriodaphnia reticulata, z veslonôžok Megacyclops viridis, Megacyclops gracilis, Eudiaptomus transylvanicus a Cryptocyclops bicolor.
Hlavný tok Dunaja
Spoločenstvá nálevníkov (Matis, Tirjakova, 1995 a, b) sa ukázali byť pomerne chudobné z hľadiska druhového zloženia i abundancie. Zastúpené boli hlavne euryékne druhy (bakteriovorné – Cyclidium glaucoma, Aspidisca cicada, A. lynceus, Glaucoma scintillans a i.). V relatívne malom počte boli zastúpené planktonické druhy. Ostatné zložky mikrozoobentosu a meiozoobentosu sa nachádzali sporadicky. Bolo to spôsobené nedostatkom vhodných substrátov, veľkou rýchlosťou prúdu, zanášaním kalom, vyplavovaním turbulenciami a nestabilitou výšky hladiny. Po záplavách Dunaja sa opakovane zisťovali inaktivácie jedincov (pravdepodobne vplyvom splavených toxických látok). V tomto období sa objavili na jednotlivých lokalitách aj zriedkavejšie sa vyskytujúce druhy napr. Ophryoglena flava, Tintinnopsis cilindrata, Stegochilum fusiforme, Frontonia anbigua, Strombidium turbo.
Permanentná fauna makrozoobentosu mala v sledovanom úseku rieky v litorálnej zóne kvalitatívne jednotný charakter (Krno et al., 1999). Medzi dominatné druhy patrili - Eunapius fragilis (Porifera), Dendrocoelum lacteum (Turbellaria), Dina punctata (Hirudinea), Ancylus fluviatilis, Lymnaea ovata, Bithynia tentaculata (Gastropoda), Dreissena polymorpha, Sphaerium corneum (Bivalvia) (Košel, 1995a). Z máloštetinavcov to boli druhy z čeľ. Naididae a Stylodrilus heringianus (Lumbriculidae), zriedkavo sa vyskytovali druhy z čeľ. Tubificidae. Významný bol aj výskyt druhu Hypania invalida (Polychaeta) a Dikerogammarus haemobaphes a Corophium curvispinum (Amphipoda). Isté rozdiely vo faune boli na profile Kľúčovec, ktorý leží už poniže spádového zlomu Dunaja (Šporka, Krno, 1995).
V taxocenózach temporárnej fauny v litoráli Dunaja dominovali Baetis fuscatus, Heptagenia sulphurea, Caenis pseudorivulorum (Ephemeroptera), Hydropsyche contubernalis, H. bulgaroromanum, Psychomyia pusilla, Brachycentrus subnubilus, Ceraclea dissimilis (Trichoptera). Oproti výskumom z 80. rokov (Krno, 1990) sme už nezaznamenali viaceré druhy podeniek - Heptagenia coerulans a rod Ecdyonurus a ďalšie druhy sa vyskytovali veľmi zriedkavo - Baetis vardarensis, Heptagenia flava, Ephemerella ignita a Potamanthus luteus. Kvantitatívne bola dunajská fauna najbohatšia pod vyústením ramien pri Gabčíkove, pričom bol výrazne bohatšie osídlený skalnatý substrát (Krno, 1995). Na ňom dominoval druh H. bulgaroromanum. Na štrkovitom substráte dominoval H. contubernalis. Celkove prevládali filtrátory (Hydropsychidae, Brachycentrus).
Mediálna časť hlavného toku Dunaja (r. km 1816), ktorú sledovala Ertlová (1968) bola osídlená hlavne máloštetinavcami (Nais elinguis, Chaetogaster crystallinus, Propappus volki, Rhynchelmis limosella a Stylodrilus heringianus), pijavicami (Erpobdella octoculata), pakomárovitými (Polypedilum sk. lateum, P. sk. scaleanum, Ablabesmyia sk. lentiginosa a Euorthocladius rivicola).
Ramená typu parapotamál
Z hľadiska štúdia mikrozoobentosu sa venovala pozornosť predovšetkým ramenám (Matis, Tirjaková, 1992; Tirjaková, 1992; Szentivány, Tirjaková, 1994). Z hľadiska štruktúry sledovaných skupín nie je možné sledované ramená hodnotiť ako celok. Spoločenstvá každého z nich, vzhľadom na svoj charakter a meniace sa podmienky, sa vyvíjali špecificky. Spoločným znakom všetkých sledovaných ramien pred rokom 1992 je dlhodobý postupný pokles hladín vody, ich premena na ramená s prúdiacou vodou len počas vyšších prietokov v Dunaji. V období po ich odrezaní od hlavného toku sa tu vyskytovali bohaté spoločenstvá typické pre stojaté vody. Ramená so stojatou vodou a ramená preplachované počas povodní mali široké druhové spektrum a abundanciu všetkých skupín mikrozoobentosu (Ciliophora, Mastigophora, Heliozoea, Amoebina).
V permanentnej faune slepých ramien, sa pôvodne vyskytovali rovnaké druhy ako v hlavnom toku - Corophium curvispinum, Dikerogammarus haemobaphes (Amphipoda), Hypania invalida (Polychaeta), Stylodrilus heringianus a rody Psammoryctides a Potamothrix (Oligochaeta) (Košel, 1995a; Krno et al., 1999). V prietočnom ramene pri Istragove dominovali, ako zistila v roku 1966 Ertlová (1970), Oligochaeta (Potamothrix moldaviensis, Tubifex tubifex, Tubifex ignotus a rod Limnodrilus) a Chironomidae (Prodiamesa olivacea, Chironomus sk. thummi, Cryptochironomus sk. defectus). Vysokú abundanciu v hlavných ramenách Bačianskej ramennej sústavy v rokoch 1976 – 1978 na štrkopiesčitom dne mediálu dosahovala Dreissena polymorpha (Bivalvia), ktorej agregácie boli vyplnené jemnými sedimentami a osídlené osobitým bentickým spoločenstvom. Druhové zloženie ako aj početnosť bentickej fauny bola vysoká (Šporka, Nagy 1998). Vysoké stavy síce spôsobovali dočasné ochudobnenie fauny, po krátkom období, cca 35 dní, sa pôvodné spoločenstvo dokázalo zotaviť.
V rámci temporárnej fauny v prietočných ramenách Lichardová (1958) pred rokom 1960 zistila viaceré taxocenózy podeniek, v ktorých sa pravidelne vyskytovali druhy – Potamathus luteus, Heptagenia sulphurea, Ecdyonurus aurantiacus, Baetis rhodani, B. fuscatus, Serratella ignita. Svedčí to o výrazne väčšej reofilnosti týchto ramien. Podobná situácia v dunajských ramenách bola s taxocenózou potočníkov (Mayer, 1935) – Rhyacophila pascoei, Agapetus sp., Hydroptila sp., Plectrocnemia sp., Neureclepsis bimaculata, Polycentropus flavomaculatus, Hydropsyche spp., Cheumatopsyche lepida, Setodes interruptus, Potamophylax latipennis, Halesus spp., Goera pilosa, Silo pallipes a Brachycentrus subnubilus. V ramenách Bačianskej ramennej sústavy v rokoch 1976 – 1978 bolo z podeniek zistených 6 taxónov a z potočníkov zástupcovia dvoch čeľadí, okrem toho z pakomárovitých bolo zistených 22 taxónov (Šporka, Nagy 1998). V rokoch 1991 – 1992 temporárna fauna prietočných ramien bola pomerne chudobná, zastúpená druhmi Cloeon dipterum, Caenis horaria, C. luctuosa (Ephemeroptera) a rodmi Ecnomus, Cyrnus, Anabolia, Athripsodes (Trichoptera). Z vážok (Majzlán, 1992) dominovali v parapotamáli druhy Calopteryx splendens a Lestes viridis. Spoločenstvo pakomárovitých sa vyznačovalo prítomnosťou druhov uprednostňujúcich tečúce typy vôd (Cricotopus bicinctus, Tanypus kraatzi) ako aj druhmi preferujúcimi pomalšie tečúce, alebo stojaté vody (Dicrotentipes spp., Polypedilum spp.) (Krno a kol., 1999).
Ramená typu plesiopotamál
V mŕtvych ramenách Dunaja sa vyskytovali pomerne stabilné spoločenstvá mikrozoobentosu (najmä na lokalitách Kráľovská lúka a Sporná Sihoť). Na lokalite Kráľovská lúka vplyvom zníženia hladiny však dochádzalo v letných mesiacoch k prehrievaniu vody, čo malo za následok zmeny v štruktúre spoločenstva hlavne nálevníkov (premnoženie niektorých druhov, napr. Coleps nolandi) (Matis, Tirjaková, 1995a,b) .
V litoráli ramien typu plesiopotamál sa vyskytovalo štrko-piesčité dno a tiež bahnité dno. Na štrko-piesčitom dne bolo druhové zloženie permanentnej fauny pestrejšie, vyskytovali sa tu okrem druhov z čeľ. Tubificidae aj druhy z čeľ. Naididae, teda druhy živiace sa nárastami, kdežto v bahnitom dne prevládali druhy z čeľ. Tubificidae, prípadne amfibiotické druhy čeľ. Enchytraeidae, veľké druhy Criodrilus lacuum a Eiseniella tetraedra (Nagy, Šporka, 1990). Druhovo chudobné boli aj ostatné skupiny, pričom prevažovali ulitníky so 14 druhmi. Abundancia väčšiny druhov bola pritom veľmi nízka, jedine kôrovec Asellus aquaticus sa v roku 1992 zistil vo zvýšenom množstve (Šporka, Krno, 1995). Z temporárnej fauny sa tu vyskytovali hlavne stagnikolné druhy vážok Sympetrum flaveolum, Lestes barbarus, Cordulia aenea ale bol tu zistený semireofilný druh Platycnemis pennipes (Majzlan, 1992). Spoločenstvo pakomárovitých bolo druhovo chudobné, pričom dominantné postavenie zaujímali pelofilné druhy Cryptochironomus defectus a Polypedilum nubeculosum (Krno et al., 1999).
V ramene Žofín v roku 1971 medzi dominantné skupiny patrili Oligochaeta a Chironomidae (Ertlová, 1973). K rovnakým záverom dospeli aj Nagy a Šporka (1990) v rokoch 1981 – 1987 na ramene pri Kráľovskej lúke, kde najvyšší počet taxónov u permanentnej fauny bol zistený v litoráli (bez zárastov a so zárastami), mediálna zóna bola osídlená len chudobne. Mediálnu zónu s bahnitými sedimentami osídľovali v prevažnej miere larvy pakomárovitých, ktoré sa druhovo početne vyskytovali aj v litorálnej zóne ramena.
Pôvodné ichtyocenózy v hlavnom toku a ramenných sústavách od ústia rieky Moravy po ústie rieky Ipľa opisuje Balon (1966), na základe prác Laboratória rybárstva v rokoch 1953–1961. Udáva existenciu 56 druhov rýb s komentárom ich výskytu. Výskumy v nasledujúcich troch desaťročiach však ukázali, že počet druhov žijúcich v tomto úseku Dunaja je výrazne vyšší. Najnovší zoznam druhov rýb slovenského úseku Dunaja obsahuje 76 položiek. Z toho 61 druhov je pôvodných, 10 (11) introdukovaných exotických a 3 (4) druhy sem invadovali z dolných úsekov Dunaja (Holčík, 2003). V porovnaní so všetkými slovenskými riekami ichtyocenóza tohoto úseku Dunaja je druhovo najbohatšia. Autor si to vysvetľuje dvomi faktormi:
1) skutočnosťou, že ide o začiatok podhorskej zóny, presnejšie, o prechod medzi podhorskou a nížinnou zónou – medzi hyporitrálom-epipotamálom a metapotamálom v zmysle klasifikácie Illies a Botosaneanu (1963);
2) zmenou spádu koryta Dunaja, ktorá zapríčinila vznik rozsiahlej vnútrozemskej delty s komplikovaným systémom ramien.
Toto spôsobuje, že na relatívne krátkom úseku Dunaja sa nachádzajú veľmi rôznorodé typy prostredia, čo podporuje vytváranie druhovo bohatého spoločenstva rýb.
Antropické zásahy v minulosti ovplyvnili ichtyocenózy v Dunaji. Výstavba vodného diela „Železné vráta“ na dolnom Dunaji ovplyvnila výskyt migrujúcich druhov. Naviac v Dunaji pribudli introdukované – allochtónne druhy. V období rokov 1970 – 1980 pri kulminácii organických a toxických polutantov došlo skoro k vymiznutiu lososovitých druhov citlivých na znečistenie, ako aj Cottus gobio, Phoxinus phoxinus, Alburnoides bipunctatus a Barbus barbus. Zlepšenie kvality vody v rokoch 1980 -1990 sa prejavilo ich opätovným výskytom. Od r.1994 sa začali objavovať aj nové druhy rýb z čeľade Gobiidae, predtým známe iba z dolných úsekov Dunaja.
Konektivita hlavného toku Dunaja s ramennými systémami priaznivo ovplyvňovala ich druhovú diverzitu. Ako konštatuje Holčík (2001), hodnoty počtu druhov, druhovej diverzity a vyrovnanosti sú v hlavnom toku vyššie a laterálne smerom od hlavného toku ku okraju vnútrozemskej delty poklesávajú. Príčinou sú rozdielne podmienky prostredia v ekosystémoch jednotlivých typov vôd vnútrozemskej delty. Nebyť vnútrozemskej delty, druhová diverzita v hlavnom toku by bola nižšia. Spôsobuje to hlavne existencia vhodných neresísk v ramenách a vhodné refúgia, hlavne počas povodní, ako aj dobrá potravinová ponuka ramien. Vyhodnotenie pôvodného stavu ichtyocenóz na monitorovacích plochách a v Dunaji pred prehradením Dunaja v roku 1992, uvádza Černý (1999).
Pri posudzovaní kvality súčasného stavu akéhokoľvek ekosystému a vývoja, akým k nemu dospel, musíme rozlišovať prírodnosť, prirodzenosť a antropicky podmienenú, alebo priam antropogénne vzniknutú umelosť. Pri posudzovaní ekosystému rieka-zaplavované územie treba okrem toho vedieť, že ide o azonálny ekologický systém, v ktorom je rozhodujúcim faktorom hydrologický režim (Lisický, 2001; Lisický, Mucha, (eds.), 2003).
Aj keď sa názory odborníkov líšia pri posudzovaní toho čo možno považovať za prírodné, teda pôvodné a človekom neovplyvnené, a do akej miery je pri antropických zásahoch potlačovaná prirodzenosť, ako aj kde leží hranica medzi prirodzeným a umelým, teda antropogénnym, keďže aj človek je v konečnom dôsledku jeden z prírodných druhov biosféry, existujú isté pragmatické, konvenčne stanovené hranice. Všeobecná zhoda panuje v tom, že ak trváme na kritériu pôvodnosti ako absencii akéhokoľvek ľudského vplyvu, potom pôvodné ekosystémy v súčasnosti prakticky neexistujú, nakoľko minimálne antropogénnymi zmenami atmosféry je priamo či nepriamo ovplyvnená celá biosféra. Naviac, antropogénne ovplyvnený je aj globálny kolobeh vody. Ak však antropický vplyv neponímame takto fundamentalisticky, potom je možné hovoriť o zvyškoch pôvodných ekosystémov v regiónoch, ktoré neboli nikdy v histórii ľudstva súčasťou ekumény, ktoré teda človek priamo nikdy neovplyvňoval, a to ani extenzívnym využívaním. Potom je možné hovoriť napríklad o zvyškoch pôvodných lesných ekosystémov v Južnej Amerike či na Sibíri (pralesoch), kým v Európe ide v najlepšom prípade o pralesovité porasty, ktoré sa spontánne vyvinuli po ukončení ľudských zásahov ako prirodzené. Ako kritérium takejto prirodzenosti možno použiť práve zachovanie základných funkčných väzieb ekosystému, ktoré zaručujú jeho spontánnu obnovu po ukončení antropického tlaku. Takéto prirodzené ekosystémy sa prirodzeným vývojom znova približujú ku kvalite prírodných, z ktorých sú odvodené, možno ich teda označiť aj ako „blízke prírodným“, prípadne aj „prírodné“, nie však ako pôvodné (Lisický, 2001).
V prípade rieky a jej ohrádzovaného záplavového územia dochádza bez ohľadu na ďalšie hospodárenie či nevyužívanie tohto územia k zásadnej zmene hydrologického režimu. Vplyv záplav sa z väčšiny pôvodnej plochy odstránil, množstvo vody však ostáva rovnaké, a preto je dynamika vodného režimu v ohrádzovanom priestore rádovo intenzívnejšia. Ohrádzované záplavové územie má potom nepôvodnú ekologickú valenciu, tak v hydrologických ako aj pedologických (a najmä pedogenetických) parametroch. Táto zmenená prirodzenosť vedie k vytvoreniu odvodeného ekosystému, ktorý je odlišný od pôvodného a ponechaný sám na seba neobnovuje kvalitu pôvodnú, ale adaptívnu. Jeho vývoj je totiž usmerňovaný obmedzením, ktoré predstavuje udržiavaná protipovodňová ochrana priľahlého územia.
Antropické zásahy do ekosystémov vedú obvykle k vytvoreniu ostrovčekov prirodzených ekosystémov, ktorých ďalší vývoj a prežívanie sú pri výraznom obmedzení pôvodnej plochy silne ovplyvňované exogénne. V prípade rieky a záplavového územia dochádza k inverzii, kedy prvotný exogénny (antropický) zásah vedie k trvalej podpore endogénneho faktora, ktorý sa po silnej redukcii pôvodnej plochy („spútaní“) stáva pre existenciu ekosystému dominantným! V ekosystéme ohrádzovaného záplavového územia sa posilňuje selekcia organizmov smerom k stenotopnosti, viazanej práve na tento typ hydrologického režimu a tvorby pôd. Ak teoreticky pripustíme návrat do stavu bez protipovodňových hrádzí, neprišlo by k samoobnovovaniu ekosystému, ktorý býva v záplavovom území po niekoľkých storočiach existencie hrádzí považovaný za blízky prírodnému či pôvodnému. Naopak ak, znovu teoreticky, pripustíme stav, keď tento dominantný faktor v území vylúčime, akokoľvek benevolentne chápaná prirodzenosť ekosystému sa stráca a jeho prežitie sa stáva závislým na lesohospodárskej pestovateľskej činnosti. Z lesa prirodzeného sa stáva les umelý, neschopný samoobnovy (Lisický, 2001).
Tieto úvahy sa týkajú nielen terestrických, ale aj veľkej časti semiakvatických spoločenstiev. Naproti tomu vodné spoločenstvá možno v regionálnom pohľade považovať za pôvodné, len čiastočne narušené prenikaním, alebo vysadením viacerých nepôvodných druhov živočíchov. Zmeny všetkých štyroch typov potamonu sa týkajú podielu ich zastúpenia vo vodnej biote vyplývajúcich z historických, najmä však recentných antropogénnych hydrogeomorfologických zmien.
Súčasná miera odvodenosti a nepôvodnosti prírodného prostredia v predmetnom území je výsledkom predovšetkým lesníckych, plavebných, vodohospodárskych ale aj poľnohospodárskych a ďalších antropických zásahov. Čím viac ideme do minulosti, tým je prírodnosť ekosystému vyššia a pre oblasť inundácie typickejšia. Z hľadiska funkčnosti dnešnej inundácie sa nemôžeme vrátiť dozadu, pred obdobie kedy boli postavené protipovodňové hrádze. Ale je možné pozmenený hydrologický režim do určitej miery a v mierke odpovedajúcej zmenenej (v našom prípade zmenšenej) inundácii, vrátiť do stavu aký bol v niektorej etape minulosti.
Funkciou dnešnej novej zúženej inundácie, je tak ako v minulosti, previesť a rozptýliť povodňovú vodu, a tak chrániť územie za protipovodňovými hrádzami i územie ležiace nižšie po toku rieky. Hydrologická funkcia inundácie ostala. V súlade s ňou je možné vytvoriť také hydrologické podmienky ktoré zachovajú základné funkčné väzby ekosystému a zabezpečia jeho spontánnu obnovu tak, aby sa prirodzeným vývojom znova priblížili ku kvalite prírodných, z ktorých sú odvodené, a aby sa tak stali aspoň blízke prírodným.
Inundácia (záplavové územie) je plochá časť terénu susediaca s riekou a pozostávajúca z aluviálnych usadenín uložených riekou. Erózno-sedimentačné procesy, premiestňovanie meandrov po prúde, vytvárajú inundáciu s meandrujúcou riekou a riečnymi ramenami. V čase povodní, keď sa rieka vylieva ponad brehy, usadzuje za brehmi a okolo riečnych ramien sedimenty. Vytvárajú sa nové ramená a staré sú zazemňované. Inundačné územie je často zaplavované a dnes siaha po ochranné protipovodňové hrádze, ktoré sú situované vo vnútri pôvodnej inundácie.
Pôvodné inundačné územie sa zmenšilo. Dlhodobá riečna erózia, úpravy riečneho koryta, včítane regulácie šírky koryta pre nízku vodu, priepichy oblúkov, uzatváranie laterálnych ramien, znížili hladinu vody v Dunaji a už dávno zhoršili ekologické podmienky inundácie. V riečnych ramenách na gabčíkovskom úseku Dunaja v období pred prehradením Dunaja sotva tiekla voda a na nagymarošskom úseku je tomu tak aj dnes, a naviac, majú mizernú kvalitu vody. Takáto situácia je popísaná v maďarskom materiáli (Úrad predsedu vlády MR, 1999) pre Nagymarošskú časť Dunaja. Podobná, ešte horšia situácia bola popísaná expertmi EU v Gabčíkovskom úseku Dunaja pred prehradením, kedy voda tiekla v skoro všetkých dunajských ramenách len 17 dní do roka a v hlavných ramenách tiekla len 3 mesiace do roka, (CEC, 1992).
Inundačné územie medzi protipovodňovými ochrannými hrádzami, v prípade stavieb Gabčíkovskej priehrady, bolo zachované počas stavebných prác v predchádzajúcom „originálnom“ stave. Lodná doprava bola preložená do derivačného kanála v dĺžke 40 km, ktorý je situovaný mimo inundačného územia. To je na Dunaji z ekologického hľadiska nová a unikátna situácia. Z hľadiska Projektu Vodného diela Gabčíkovo, hlavnou funkciou inundácie je previesť povodňovú vodu, ktorá nemôže pretiecť derivačným kanálom (prietok v Dunaji pri Bratislave mínus prietok, ktorý pretečie derivačným kanálom). Druhou funkciou inundácie je funkcia prírodného poldra, čo znamená dočasne uskladniť vody povodňového maxima medzi ochrannými hrádzami, a tak znížiť maximálne povodňové prietoky smerom po prúde od Gabčíkova smerom na Budapešť. Z ekologického hľadiska to znamená, že toto územie, medzi Čunovom a Sapom, má byť ekologicky chápané a podporované ako pravidelne zaplavované územie so všetkými dôsledkami. To znamená, že má mať dostatočnú kapacitu previesť povodňovú vodu a dočasne uskladniť časť maximálnych povodňových prietokov. V čase medzi povodňami a tiež počas malých prietokov vody v Dunaji, v riečnych ramenách je a aj má byť tečúca voda, zabezpečená odbernými zariadeniami postavenými v rámci Projektu a priamym prepojením so starým korytom Dunaja. Že je to možné, dokumentujú technické opatrenia – zavzdutie vody – realizované v rámci Dohody z roku 1995. Samočistiace procesy vo vode sú v riečnych ramenách aktívne len ak je v nich voda a to predovšetkým prúdiaca voda. Naviac, pre inundáciu typické biotopy majú byť podporované kontinuálnym napájaním ramien vodou a simuláciou povodní niekoľkokrát do roka, v súlade s reálnymi prietokmi vody v Dunaji nad týmto územím. Takýto základný vodný režim má byť podporovaný niektorými ďalšími technickými opatreniami.
Cieľom ďalších opatrení a návrhov optimalizácie (Lisický, Mucha, (eds.), 2003) a manažmentu má byť obnovenie integrity ekosystému v zaplavovanom území medzi pôvodnými protipovodňovými hrádzami (v inundácii). Z toho dôvodu sa v ňom treba zamerať na podporu rozhodujúcich ekologických procesov. Nejde pritom o zabezpečenie optimálnych životných podmienok pre čo najväčší počet druhov, ale o zachovanie prirodzenej biodiverzity, udržanie amplitúdy a frekvencie zmien (dynamická rovnováha) a zachovanie reparačných a regeneračných procesov.
Optimalizácia by mala viesť k návratu k prírodnosti (nie pôvodnosti), t. j. k zabezpečeniu resp. vytvoreniu podmienok pre prirodzené uplatňovanie ekologických zákonitostí a autoreguláciu celého ekosystému ako komplexu azonálnych ekosystémov. Ak vychádzame zo základnej predstavy vnútrozemskej delty tvorenej katenou spoločenstiev od akvatických (eupotamál až plesiopotamál) cez semiakvatické litorálne a prechodné až po terestrické, určujúcim vlhkostným gradientom, tomu zodpovedajúcou hladinou podzemnej vody a cyklickými disturbanciami v podobe záplav, potom cieľom by malo byť dosiahnutie stavu, v ktorom by sa po umelom vytvorení podmienok mohlo realizovať samoregulačné fungovanie a v istom zmysle sekundárna sukcesia tých spoločenstiev, ktoré sa od prirodzeného stavu rôznymi zásahmi vzdialili. Je zrejmé, že dôsledkom takéhoto vývoja môže lokálne dôjsť aj k zmenám až zániku existujúcich spoločenstiev, podmienky by však mali byť nastavené tak, aby prirodzenou sukcesiou (s možnosťou usmernenia vývoja ekosystému v prvom období) tieto vznikli na iných vhodných miestach. Pri dôslednom realizovaní tejto predstavy musíme pripustiť aj možnosť vzniku spoločenstiev zo suchšej oblasti vlhkostného gradientu (tvrdé lužné lesy, dunajská lesostep, ktoré tu v minulosti hojne existovali), ktoré sa v tejto oblasti v súčasnosti začínajú formovať v drénovanej časti inundácie.
Potenciálny stav spoločenstiev nemôže byť pritom chápaný ako statický stav, ale ako dynamické oscilácie medzi dvoma idealizovanými prirodzenými stavmi (extrémne mokrým s opakovanými silnými disturbanciami a relatívne suchším a stabilizovanejším). Tento potenciálny stav pritom nemôže byť považovaný za nemennú charakteristiku konkrétneho miesta, ale iba za medze, cez ktoré by sa stav konkrétneho spoločenstva nemal dlhodobo vychýliť. Znamená to, že systém sa môže dynamicky meniť nielen v čase, ale aj v priestore.
Tento stav nemusí automaticky znamenať zachovanie všetkých, v súčasnosti sa tu vyskytujúcich druhov a teda zachovanie úplnej súčasnej druhovej diverzity. Tá je totiž v súčasnosti tvorená aj celým radom ekologicky inváznych druhov so širšou ekologickou toleranciou, ktorých sekundárny prienik je podmienený práve zmenenými podmienkami. Práve pre potlačenie inváznych druhov rastlín, do čias kým to neprevezme autoregulácia, je potrebná možnosť usmerňovania prebiehajúcej renaturácie. K takémuto usmerňovaniu je potrebný účelovo zameraný monitoring, metodika jeho vyhodnotenia a nástroje na usmerňovanie.
Pre podrobnejšiu záverečnú diskusiu o ďalšom postupe Lisický, Mucha (2003) a celý rad autorov v citácii autorov monografie „Optimalizácia vodného režimu ramennej sústavy v úseku Dunaja Dobrohošť – Sap z hľadiska prírodného prostredia“ odporúčajú návrhy patriace do skupiny – nový eupotamál. Prednosť má komplexné slovensko-maďarské riešenie, ktoré by obnovilo jednotný ekosystém v tomto úseku Dunaja. Z praktického hľadiska zabezpečenia prevedenia povodňových prietokov a cieľov ekológie vytvoriť nový eupotamál sa zdá byť východiskom vytvorenie nového eupotamálu rieky s využitím pretekaných prehrádzok v miestach, ktoré križujú staré koryto Dunaja a v miestach, kde je treba znížiť drenážny účinok starého koryta Dunaja. Takéto riešenie zachová prevádzanie povodňových prietokov starým korytom Dunaja, zníži drenážny účinok starého koryta a s malým (premenlivým) prietokom starým korytom bude fungovať pokiaľ sa nesfunkční eupotamál v nosných ramenách pôvodnej ramennej sústave a ak sa to ukáže vhodné tak aj naďalej. Počas povodne, staré koryto prevezme funkciu prevedenia prietokov, ktoré neprejdú novým eupotamálom a zaplavenou inundáciou. Nový eupotamál by mal byť taký, aby na väčšine svojej dĺžky nepotreboval údržbu svojho koryta a opevňovanie brehov.
Ak prijmeme zásadu, že úpravy by mali byť nenásilné, nemali by sa realizovať veľkoplošne v inundačnom území, nemali by byť nákladné, nemali by obsahovať veľké objemy zemných prác a mali by konvergovať k prírodným procesom, potom existuje určitá kombinácia návrhov a určitá postupnosť prác. To by malo aj výhodu možnosti úprav priebehu prác na základe monitorovania prírodného prostredia. Tento postup môže byť v čase nasledujúci (Lisický, Mucha, (eds.), 2003):
vybudovanie niekoľkých nízkych prehrádzok (nie však pevných hatí), umožňujúcich na niekoľkých miestach prepojenie ramenných sústav so starým korytom Dunaja,
zabezpečenie prirodzeného delenia vody medzi staré koryto Dunaja a prepojené ramená,
optimalizovanie prietokov do ramien vo vzťahu k prehrádzkam a k dynamike vody v ramenách,
úpravy vedúce k obnoveniu prírodných procesov autoregulačného ekosystému, vrátane dynamiky prietokov a hladín povrchových a podzemných vôd, erózie a sedimentácie, meandrovania v inundačnom priestore, vzájomného prepojenia ramien.
monitorovanie funkčnosti, na začiatku niekoľkých vybudovaných (2-3) nízkych prehrádzok v starom koryte Dunaja a na základe výsledkov navrhnutie ďalších úprav, napríklad spevnenia brodov, prepojenia medzi ramenami, zmeny výšky hladín a i. Postupné budovanie ďalších prehrádzok v Dunaji.
Experti, ktorí pripravili prácu Lisický, Mucha, (eds.), (2003), sa prikláňajú k myšlienke postupného riadeného (technickými prácami a vodohospodárskym manažmentom usmerňovaného) uvoľňovania riečnej dynamiky v predmetnom území podľa v závere navrhnutého scenára a k zásade postupného adaptívneho rozhodovania smerujúceho k postupnej optimalizácii vodného režimu v nasledujúcich desaťročiach, ktorej cieľom sú základné myšlienky vyjadrené v predchádzajúcich odsekoch. Pritom odporúčajú realizovať v ramennej sústave čo najmenej technických prác a ponechať pretvorenia nového eupotamálu prírode a prírodným procesom.
Áč, P., Bohuš, M., Novacký, P., 1996: Výskyt a priestorové vzťahy akvatických druhov vtákov v inundácii Dunaja a vo vodnom diele Gabčíkovo. Acta Environmentalica Universitatis Comenianae, 6, s. 129 – 136.
Adam, Z., Dlabač, M., 1961: Nové poznatky o tektonice Podunajské nížiny. Věst. ÚÚG, 36/3 s. 189-198.
Antonič, M., Jacko, R., 1966: Hydrochémia československého úseku Dunaja a jeho prítokov. In Mucha V., Dub O. et al., 1966: Limnológia československého úseku Dunaja, SAV, Bratislava, s. 105-168.
Archívne materiály Geofondu v Bratislave.
Balát, F., 1963: Ptačí fauna Žitného ostrova. Biologické práce, 9(7), s. 1-84.
Baláži, P., Matis, D., 2002: The species diversity, seasonal dynamics and trophical relations of the pelagic ciliated protozoa in the selected rivers of Slovakia. Ekológia (Bratislava), 21(1), s. 3-14.
Baláži, P., Tirjaková, E., 1999: New records of Suctoria (Ciliophora) from Slovakia. Biológia (Bratislava), 55(2), s. 149-150.
Balthasar, V., 1936: Limnologické výskumy v slovenských vodách. Bratislava, SUS, 75 s.
Balthasar, V., 1938: Další příspěvek k entomologickému výskumu Slovenska. Entomol. listy, 2, s. 122-123.
Balon, E., K., 1966: Ichtyofauna československého úseku Dunaja. In Mucha, V. - Dub, O. et al., 1966: Limnológia Československého úseku Dunaja, SAV, s. 270-323.
Bertová, L., et al., 1986: Floristic characteristic from standpoint of genofond rare and threated species. In: Vegetation bases to assanation and reconstruction of the Hrušov reservoir territory, I., II., ÚEBE SAV Bratislava, rukopis.
Biely, A., (ed), 1996: Geologická mapa Slovenska, Vysvetlivky ku geologickej mape Slovenska, Vyd. Dionýza Štúta, Bratislava, 77 s., mapa.
Brtek, J., Rotschein, J., 1964: Ein Beitrag zur Kenntnis der Hydrofauna und des Reinheitszustandes des Tschechoslovakischen Abschnites der Donau. Biol. Práce, 10 (5), s. 1-64.
Bulánková, E., 1995: Nabinae (Nabidae, Heteroptera) Podunajskej nížiny. Entomofauna carpathica, 7, s. 83-92.
Bulánková, E., 1999: Zmeny v druhovom zložení vážok (Odonata) Podunajskej roviny za posledných tridsať rokov. Entomofauna Carpathica, 11/1, s. 1-5.
Bulánková, E., 2001: Vážky ako indikátory zmien životného prostredia v oblasti VD Gabčíkovo. Acta Facultatis Rerum Naturalium Ostraviensis 200, Biológia – Ekológia, 8, s. 88-92.
Bulánková, E., 2001: Epifauna v oblasti VD Gabčíkovo. Vážky (Insecta: Odonata). In Optimalizácia 2003.
Bulánková, E., Halgoš, J., 1995: Výsledky monitoringu vybraných skupín hmyzu (Odonata, Culicidae) stojatých vôd v oblasti VDG. Súhrn referátov z konferencie „Výsledky a skúsenosti z monitorovania bioty územia ovplyvneného vodným dielom Gabčíkovo“, 15. 16. 11. 1994., Bratislava, s. 418-425.
Bulánková,E., Halgoš, J., 1997: The influence of flowing water on the occurrence of dragonflies and blackflies in the Danube and the Morava river basin. Limnologische Berichte, Donau 1997, Band I, 32. Konferenz der IAD, Wien, s. 293-297.
Cec, 1992: Commission of the European Community, Czech and Slovak Federative Republic, Republic of Hungary 1992. Working Group of Independent Experts on Variant C of the Gabčíkovo-Nagymaros Project, Working Group Report, Budapest, Nov. 23, 1992.
Čarnogurský, J., Krumpálová, Z., Kalúz, S., Wirthová, M., 1994: Soil Arthropods of Forest and Adjacent Agrocoenoses in Certain Localities of the Danube Region in Southwestern Slovakia. Biologia (Bratislava), 49, s. 173-183.
Čejka, T., 2000: Monitoring vplyvu prevádzky VD Gabčíkovo na biotu (časť Gastropoda). Záverečná správa za rok 2000. Ms. depon. in ÚZ SAV a GWC, 6 s.
Černý, J., 1999: Monitoring of ichthyocoenoses in the Slovak stretch of the Danube and in its side armes. p. 201 - 216. In: Mucha, I. (ed.): Gabčíkovo part of the hydroelectric power project - Environmenal impact review. Faculty of Natural Sciences, Comenius University, Bratislava. 399 s.
DANREG, 2000, Császár (ed.): Danube Region Environmental Geology Programme, DANREG Explanatory Notes, Jahrbuch, Geologishe Bundesanstalt, Wien, 606 s.
David, S., 1994: Nové nálezy vzácnych a ohrožených druhů vážek (Insecta: Odonata) jihozápadního Slovenska. Acta musei Tekovensis, Levice, 2, s. 81-90.
Directive 2000/60/EC of the European Parliament and of the Council, aimed at maintaining and improving the aquatic surface and ground water environment.
Dosztányi, I., 1988: Gabčíkovo-Nagymaros, Environment and river dams, Aqua Kiadó, Budapest, 36 s.
Dub, O., Szolgay, J., 1966: Hydrologia a hydrogeológia československého úseku Dunaja. In: Mucha, V., Dub, O. (eds.) Limnológia československého úseku Dunaja. VSAV, Bratislava, s. 9-60.
Ertl, M., 1970: Zunahme der Abundanz der Periphyton-Mikrofauna aus der Donau bei Besiedlung der Substrate. Biologické práce SAV, 16, 3, s. 1-104.
Ertl, M., 1976: Primárna produkcia planktónu v strednom toku Dunaja. Biologické práce SAV, 22, 4, s. 81-140.
Ertlová, E., 1963: Zoobentos mŕtvych ramien Dunaja. Biológia 18, s. 743-755.
Ertlová, E., 1968: Die Mengen des Zoobenthos in den Schottern des Donaumedials. Arch. Hydrobiol. 34, s. 321-330.
Ertlová, E., 1970: The abundance of the bottom fauna in an arm of the river Danube with flowing water. Biologia 25, s. 521-526.
Ertlová, E., 1973: Poznámky o abundancii a biomase makrozoobentozu dunajského ramena ležiaceho neďaleko obce Vojka. Zoologia 19, s. 79-85.
Ertlová, E., 1987: Chironomids (Chiromnomidae, Diptera) in the Danube et Bratislava. Acta F.R.N. Univ. Comen.- Zoologia 29, s. 67-78.
Fink, J., 1966: Die Paläogeographie der Donau. In Limnologie der Donau, (Hrsg. R. Liepold) Liefg.2. Stuttgart, s. 1-50.
Flasarová, M., 1999: Bericht über Isopoden (Asellota et Oniscidea) im slowakischen Donaugebiet. Acta Mus. Nat. Pragae, Series B, Historia Naturalis, 54 (1998), s. 61-78.
FNSCU, 1995: Gabčíkovo Part of the Hydroelectric Power Project, Environmental Impact Review, (Evaluation Based on Two Year Monitoring), (Mucha, I., sc. Ed.), Faculty of Natural Sciences, Comenius University Bratislava, Slovakia and Plenipotentiary of the Slovak Republic for Construction and Operation of Gabčíkovo-Nagymaros Hydropower Scheme, Bratislava, 384 s.
FNSCU, 1999: Gabčíkovo Part of the Hydroelectric Power Project, Environmental Impact Review, (Evaluation Based on Six Year Monitoring), (Mucha, I., sc. Ed.), Faculty of Natural Sciences, Comenius University Bratislava, Slovakia and Plenipotentiary of the Slovak Republic for Construction and Operation of Gabčíkovo-Nagymaros Hydropower Scheme, Bratislava, 399 s.
Franko, O., Remšík A., Fendek, M., Bodiš, D., 1984: Geotermálna energia centrálnej depresie podunajskej panvy – prognózne zásoby. Čiastková záverečná správa GÚDŠ, Bratislava.
Franko, O., Bodiš, D., Fendek, M., Remšík A., Jančí, J., Král, M., 1989: Methods of research and evaluation of geothermal resources in pore environment of Panonian Basin. Západné Karpaty, sér. Hydrogeol a inž. Geol. 8, GÚDŠ, Bratislava, s. 165 – 192.
Gašparík, J., 1979: Paleogeografické mapy Slovenska – neogén, Geologický ústav Dionýza Štúra, Bratislava, 33 s., + mapy.
Gulička, J., 1957: Kvalitatívno - kvantitatívny rozbor pôdnej fauny Čierneho lesa (Ostrov), (Myriapoda, Isopoda, Opiliones). Acta F.R.N. - Univ. Comenianae, Zoologia, 3, s. 119-139.
Halgoš, J., 1981: Sezónna dynamika komárov (Diptera, Culicidae) v Bratislave. Referáty vedecko-pracovnej konferencie "Provázkove dni" v Komárne, s. 18-21.
Halgoš, J., 1984: Culicidae. In Čepelák, J., (ed.): Diptera Slovenska I. Veda SAV (Bratislava), s. 48-61.
Halgoš, J., 1995: The monitoring of mosquitoes (Diptera, Culicidae) in the area affected by the Gabčíkovo hydroeletric power structures. In Mucha, I. (ed.): Gabčíkovo part of the hydroelectric power - enviromental impact review, Faculty of Natural Sciences, Comenius University, Bratislava, Slovakia, s. 311-314.
Harčár, J., Priechodská, Z., 1988: Geologická mapa Podunajskej nížiny, SV časť, 1:5000, GÚDŠ Bratislava.
Hejný, S., 1960: Ökologische Charakteristik der Wasser und Sumpflanzen in den slowakischen Tiefebene (Donau – und Theissgebiet). SAV Bratislava, 448 s.
Holčík, J., 2001: Ryby slovenského úseku Dunaja. In Optimalizácia 2003.
Holčík, J., 2003: Past and prresent fish fauna and fisheries in the Slovak segment of the Danube river: A review. Ann. Limnol. – Int. J. Limnology.
Holčík, J., Bastl, I., Ertl, M., Vranovský, M., 1981: Hydrobiology and ichthyology of the Czechoslovak Danube in relation to predicted changes after the construction of the Gabčíkovo-Nagymaros river barrage system. Práce Lab. rybár. hydrobiol., 3, s. 19-158.
Hovorka, D., Michalík, J., 2001: O čom hovoria naše vrchy – Geológia pre každého, VEDA, Bratislava, 276 s.
Hrabě, S., 1941. K poznání dunajských Oligochet. Acta Soc. Scient. Natur. Moravicae, 137, s. 1-37.
Cheben, I., Hajnalová, E., Čejka, T., 2001: Waldvegetation des Unterlaufes der Žitava im Neolithikum. Pravěk, suppl. 8, s. 40-52.
Illies, J., Botosaneanu, L., 1963: Problemes et méthodes de la classification de la zonation écologique des eaux courrantes, considerés surtout du point de vue faunistique. Mitt. Int. Verein. Theor. Angew. Limnologie 12, 57 s.
Illyová, M., 1995: Výsledky monitoringu perloočiek (Cladocera) na území vnútrozemskej delty Dunaja. In Svobodová, A., Lisický, M.J. (eds.).: Výsledky a skúsenosti z monitorovania bioty územia ovplyvneného VD Gabčíkovo. Ústav zoológie SAV, Bratislava, s. 86-93.
Janáček, J., 1966: Vodné dielo Dunaj – výzkum tektoniky. GÚDŠ Bratislava, správa.
Janáček, J., 1967: Výskum tektoniky južnej časti Podunajskej nížiny s ohľadom na výstavbu vodného diela Dunaj. Záverečná správa, GÚDŠ.
Jedlička, L., Országh, I., Čejka, T., Darolová, A., Kulfan, M., Mikulíček, P., Šustek, Z., Žiak, D., 1999: Terrestrial fauna. In: Mucha, I. (ed.): Gabčíkovo Part of the Hydroelectric Power Project - Environmental Impact Review. Faculty of Natural Sciences, Comenius University, Bratislava, s. 323-381.
Joint Annual Report of environmental monitoring in 1995 according to the "Agreement between the Government of the Slovak Republic and the Government of Hungary about Certain Temporary Measures and Discharges to the Danube and Mosoni Danube", signed April 19, 1995, submitted jointly by the Slovak and Hungarian sides including Minutes from meetings of 29 May 1995, 29 November 1995 and 25 March 1996. Nominated Monitoring Agents of the Government of the Slovak Republic and the Republic of Hungary.
Joint Annual Report of environmental monitoring in 1995 according to the "Agreement between the Government of the Slovak Republic and the Government of Hungary about Certain Temporary Measures and Discharges to the Danube and Mosoni Danube", signed April 19, 1995, Exchanged data and minutes from negotiations. Nominated Monitoring Agents of the Government of the Slovak Republic and the Republic of Hungary.
Joint Annual Report of the environment monitoring in 1996, according to the “Agreement between the Government of the Slovak Republic and the Government of Hungary about Certain Temporary Measures and Discharges to the Danube and Mosoni Danube, Signed April 19, 1995. Nominated Monitoring Agents of the Government of the Slovak Republic and the Republic of Hungary.
Joint Annual Report of the environment monitoring in 1997, according to the “Agreement between the Government of the Slovak Republic and the Government of Hungary about Certain Temporary Measures and Discharges to the Danube and Mosoni Danube, Signed April 19, 1995. Nominated Monitoring Agents of the Government of the Slovak Republic and the Republic of Hungary.
Joint Annual Report on the environment monitoring in 1998 according to the "Agreement between the Government of the Slovak Republic and the Government of Hungary about Certain Temporary Measures and Discharges to the Danube and Mosoni Danube", signed April 19, 1995. Nominated Monitoring Agents of the Government of the Slovak Republic and the Republic of Hungary.
Joint Annual Report on the environment monitoring in 1999 according to the "Agreement between the Government of the Slovak Republic and the Government of Hungary about Certain Temporary Measures and Discharges to the Danube and Mosoni Danube", signed April 19, 1995. Nominated Monitoring Agents of the Government of the Slovak Republic and the Republic of Hungary.
Joint Annual Report on the environment monitoring in 2000 according to the "Agreement between the Government of the Slovak Republic and the Government of Hungary about Certain Temporary Measures and Discharges to the Danube and Mosoni Danube", signed April 19, 1995. Nominated Monitoring Agents of the Government of the Slovak Republic and the Republic of Hungary.
Joint Annual Report on the environment monitoring in 2001 according to the "Agreement between the Government of the Slovak Republic and the Government of Hungary about Certain Temporary Measures and Discharges to the Danube and Mosoni Danube", signed April 19, 1995. Nominated Monitoring Agents of the Government of the Slovak Republic and the Republic of Hungary.
Joint Annual Report on the environment monitoring in 2002 according to the "Agreement between the Government of the Slovak Republic and the Government of Hungary about Certain Temporary Measures and Discharges to the Danube and Mosoni Danube", signed April 19, 1995. Nominated Monitoring Agents of the Government of the Slovak Republic and the Republic of Hungary.
Joint Annual Report on the environment monitoring in 2003 according to the "Agreement between the Government of the Slovak Republic and the Government of Hungary about Certain Temporary Measures and Discharges to the Danube and Mosoni Danube", signed April 19, 1995. Nominated Monitoring Agents of the Government of the Slovak Republic and the Republic of Hungary.
Jurko, A., 1958: Pôdne ekologické pomery a lesné spoločenstvá Podunajskej nížiny. SAV, Bratislava, 264 s.
Kalivodová, E., Darolová, A., 1998: Vtáky slovenského úseku Dunaja a Žitného ostrova. Edícia Biosféra, Vol. A1, Biosféra, Bratislava, 97 s.
Kodada, J., Majzlan, O., 1991: Spoločenstvá chrobákov (Coleoptera) Klátovského ramena pri Jahodnej (juhozápadné Slovensko). Entomol. Probl. (Bratislava), 21, s. 31-52.
Košel, V., 1995a: Ripal macrozoobenthos of the Danube before and after putting hydropower structures Gabčikovo into operation. Výsledky a skúsenosti z monitorovania bioty územia ovplyvneného vodným dielom Gabčíkovo, Ústav zoológie a ekosozológie SAV, Bratislava, s. 123-131.
Košel, V., 1995b: Permanent macrozoobenthos in Danube area before and during the operation of the Gabčíkovo. In: Mucha, I. (ed.): Gabčíkovo Part of the Hydroelectric Power Project - Environmental Impact Review. Faculty of Natural Sciences, Comenius University, Bratislava, s. 233-238.
Kováč, M., 2000: Geodynamický, paleogeografický a štruktúrny vývoj Karpatsko-panónskeho regiónu v moicéne: Nový pohľad na neogénne panvy Slovenska. VEDA, Bratislava, 202 s.
Krno, I., 1990: Investigations of Mayflies (Ephemeroptera) and Stoneflies (Plecoptera) of the Danube in the region of the Gabčíkovo Barrage. Acta F.R.N.Univ.Comen.- Zool., 33, s. 19 - 30.
Krno, I., 1995: The changes in the taxocoenoses structure of mayflies (Ephemeroptera) and caddisflies (Trichoptera) of the river Danube and the surrounding stagnant waters. In Mucha, I. (ed.): Gabčíkovo part of the hydroelectric power project - enviromental impact rewiev, Fac. Nat. Scien, Comenius University, Bratislava, s. 301-306.
Krno, I., Šporka, F., Matis, D., Tirjaková, E., Halgoš, J., Košel, V., Bulánková, E., Illéšová, D. 1999: Development of zoobenthos in the Slovak Danube inundation area after the Gabčíkovo hydropower structures began operating. In Mucha, I. (ed.): Gabčíkovo part of the hydroelectric power project - Environmental impact review. Faculty of Natural Sciences, Comenius University, Bratislava, s. 175-200.
Kubalová, S., 2001: Močiarna vegetácia stojatých a tečúcich vôd. (Expertízne vyjadrenie k optimalizácii vodného režimu v inundácii Dunaja). In Optimalizácia 2003.
Landa, V., 1969: Jepice - Ephemeroptera. Fauna ČSSR, ČSAV, Praha. 352 s.
Lászlóffy, W., (sc.ed.), 1965: Az 1954. évi Dunai árvíznek szentelt szám, Vízügyi Közlemények az Országos Vízügyi Főigazgatóság kiadványa, 1955. év 1-2. füzet.
Lichardová, E., 1958: Príspevok k poznaniu jednodňoviek (Ephemeroptera) ramien Dunaja a periodických mlák na Žitnom ostrove. Biológia, 13, s. 129-133.
Lisický M., J., 2001: Expertné vyjadrenie k optimalizácii vodného režimu ramennej sústavy z hľadiska prírodného prostredia. In Lisický, M., J., Mucha I., (eds.), 2003: Optimalizácia vodného režimu ramennej sústavy v úseku Dunaja Dobrohošť – Sap z hľadiska orírodného prostredia, Prírodovedecká fakulta Univ. Komenského v Bratislave, Monografia, 205 s.
Lisický M., J., Mucha, I., (eds.), 2003: Optimalizácia vodného režimu ramennej sústavy v úseku Dunaja Dobrohošť – Sap z hľadiska prírodného prostredia, Prírodovedecká fakulta Univ. Komenského v Bratislave, Monografia, 205 s.
Ložek, V., 1955: Zpráva o malakozoologickém výzkumu Veľkého Žitného ostrova v roce 1953. Práce II. sekcie SAV, séria biol., zv. 1., zošit 6, 32 s.
Ložek, V., 1964: Quartärmollusken der Tschechoslowakei. Rozpr. Ústř. Úst. Geol., Vol. 31, 374 s.
Maglay, J., (ed.), 1999: Neotektonická mapa Slovenska, Vysvetlivky k neotektonickej mape Slovenska 1:500000, Geol. Služba SR, Bratislava, 48 s., mapa.
Majzlan, O., 1988: Spoločenstvá chrobákov (Coleoptera) Číčova a blízkeho okolia. Entomol. Probl. (Bratislava), 18, s. 133-170.
Majzlan, O., 1990: Spoločenstvá významných fytofágnych koleopter lúčnych a lesných biotopov. Acta F.R.N. Univ. Comen. – Formatio et Protectio Naturae, 12, s. 75-93.
Majzlan, O., Rychlík, I., 1982: Chrobáky (Coleoptera) v dosahu riečneho toku Dunaja pri Bratislave. Entomol. Probl. (Bratislava), 17, s. 33-81.
Matis, D., 2001: Ekologické dopady výstavby Dunajského vodného diela. In Optimalizácia 2003.
Matis, D., Tirjaková, E. 1992: Nálevníky (Ciliophora) mŕtveho ramena Dunaja v Čičove. Acta F.R.N.Univ. Comen. Formatio et Protectio Naturae, 16, s. 49 – 56.
Matis, D., Tirjaková, E. 1995a: Zmeny spoločenstiev nálevníkov (Ciliophora) riečneho systému Dunaja v rokoch 1989 – 1994. In: Výsledky a skúsenosti z monitorovania bioty územia ovplyvneného vodným dielom Gabčíkovo. ÚZE SAV, Bratislava, s. 94-103.
Matis, D., Tirjaková, E. 1995b: The impact of the hydroelectric power structures Gabčíkovo on the ciliata (Ciliophora) communities. In Mucha, I., (ed.): Gabčíkovo part of the hydroelectric power project – environmental impact review. Faculty of Natural Sciences Commenius University, Bratislava, Slovakia, s. 241 – 250.
Mayer, K., 1935: Příspěvek k poznaní chrostíkú okolí Čilistova na Dunaji. Sborník Přír. Klubu v Brne, 17, s.73-78.
Mazúr, E., et al., 1980: Atlas SSR. Tématická skupina V. Ovzdušie a vodstvo. SAV a Slov. Úrad geodézie a kartografie, Bratislava.
Michalík, J., Reháková, D., Kováč, M., Soták, J., Baráth, I., 1999: Geológia stratigrafických sekvencií, VEDA, SAV, Bratislava, 233 s.
Mucha, I., Banský, Ľ., Hlavatý, Z., 2001: Optimalizácia vodného režimu ramennej sústavy z hľadiska prírodného prostredia. Konzultačná skupina Podzemná Voda, s.r.o., Bratislava, 20 s.
Mucha, I., et al., 1999: Návšteva oblasti Vodného diela Gabčíkovo, Splnomocnenec vlády SR pre výstavbu a prevádzku Sústavy vodných diel Gabčíkovo-Nagymaros, Konzultačná skupina Podzemná Voda, s.r.o., Bratislava, 103 s.
Mucha, I., Šestakov, V., M., 1987: Hydraulika podzemných vôd. ALFA, Bratislava, 343 s.
Mucha V., Dub O., (eds.), 1966: Limnológia československého úseku Dunaja, SAV, Bratislava, 327 s.
Mucha I. et al.,1991: Databázové spracovanie hydrogeologických parametrov pre model prúdenia podzemných vôd Podunajskej nížiny, príprava dát pre program PHARE EC/WAT/1, október 1991, Univerzita Komenského, Prírodovedecká fakulta, Bratislava, Konzultačná skupina Podzemná Voda, s.r.o., Bratislava.
Nagy, Š., Šporka, F., 1990: Makrozoobentos dunajského ramena typu plesiopotamon a jeho zmeny pod vplyvom umelého zarybnenia. Biológia (Bratislava) 45, s. 781-790.
Neštický, Š., Varga, L., 2001: Optimalizácia vodného režimu ramennej sústavy z hľadiska lesného hospodárstva. Expertízne vyjadrenie k optimalizácii vodného režimu v inundácii. In Optimalizácia 2003.
Optimalizácia, 2003: Optimalizácia vodného režimu ramennej sústavy v úseku Dunaja Dobrohošť – Sap z hľadiska prírodného prostredia, Lisický M., J., Mucha, I., (eds.), Prírodovedecká fakulta Univ. Komenského v Bratislave, Monografia, 205 s.
Oťaheľová, H., 2001: Expertízne vyjadrenie k optimalizácii vodného režimu v inundácii. Makrofytná vegetácia stojatých a tečúcich vôd. In Optimalizácia 2003.
Pišút, P., 2000: Vývoj koryta Dunaja na území Bratislavy v 18.-20. storočí podľa historických máp. Doktorand. diz. práca, Ms. depon. in ÚZ SAV, Bratislava, 155 s.
Phare, 1995: Danubian Lowland – Ground Water Model, PHARE Project No. PHARE/EC/WAT/1, Ministry of Environment, Slovak Republic, Commission of the European Communities, December 1995.
Pomichal, R., 1984: Príspevok k faune potočníkov Podunajskej nížiny. Spravodajca Žitno-ostrovského múzea, 8, s. 67-80.
Pospišil, P., Melioris, L., 1981: Poznámky k hydrogeologickému charakteru podložia a výplne Gabčíkovskej prepadliny. Acta F.R.N. Univ. Comen. Formatio et protectio naturae VII., Bratislava, s. 15-25.
Pospišil, P., Vass, D., Melioris, L., Repka, T., 1978: Neotektonická stavba Žitného ostrova a priľahlého územia Podunajskej nížiny. Minerália slovaca, 10, 5, Bratislava, s. 443- 456.
Priechodská, Z., Vass, D., 1986: Geológia neogénu centrálnej depresie v Podunajskej nížine. Regionálna geológia Západných Karpát, Správy o výskumoch Geol. Úst. D. Štúra 21, Bratislava, s. 105 – 111.
Raušer, J., 1957: K poznání dunajských pošvatek (Plecoptera). Zoologické listy, s. 257-282.
Rothschein, J., Hanzlíková, G., 1966: Saprobiológia československého úseku Dunaja. In Mucha V., Dub O., Limnológia československého úseku Dunaja, SAV, Bratislava, s. 218-269.
Rovný, B., Cambel, B., Čejka, B., Svobodová, A., Uherčíková, E., 1996: Doterajšie výsledky a skúsenosti s účelovým monitoringom bioty pre potreby vodného diela Gabčíkovov, s. 132-139. In: Eliáš, P., (ed.): Monitorovanie bioty na území Slovenskej republiky, SEKOS, Bratislava.
Russev, B., 1979: Gegenwartige Kenntnisse über die Arten-zusammensetzung des Zoobenthos der Donau. 19. Internationale Arbeitsgemeinschaft Donauforschung, Sofia, s. 306-336.
Rybanič, R., 1999: Birds. In Mucha I. (ed.): Gabčíkovo Part of the Hydroelectric Power Project – Environmental Impact Review. Faculty of Natural Sciences, Comenius University, Bratislava, s. 383-399.
Scharek, P., ed., 1991: The Geological Map Series of the Little Hungarian Plain, Mosonmagyaróvár, Győr – Észak, Hungarian Geological Institute).
Spoločný zmluvný projekt 1966: SVD G-N – spoločný zmluvný projekt, Pozdĺžny profil Dunaja, OVIBER, VIZITERV Budapešť, VVIP, Hydroconsult Bratislava.
Svobodová, A., 1994: Evolution of arm system and their functional typology (example of the slovak-hungarian Danube River reach). Ekológia, Bratislava 13, 4, s. 369-383.
Szentivány, M., Tirjaková, E., 1994: The structure and dynamics of the community Ciliophora in benthos of the Karlova Ves (Bratislava) branch of the Danube. Acta Zool. Univ. Comen., 38, s. 83-102.
Šíbl, J., Seginková, A., Bulánková, E., 2001: Príspevok k poznaniu fauny vážok (Odonata) Podunajskej roviny. Entomofauna Carpathica, 13, s. 68-71.
Šiška, B., 1983: Cenózy bzdôch (Heteroptera) v korunách vybraných druhov drevín Podunajska. Diplomová práca, Prírodovedecká fakulta UK Bratislava, 83 s.
Šomšák, L., 1999: Flora and vegetation conditions of floodplain ecosystems. In Mucha I. (ed.): Gabčíkovo Part of the Hydroelectric Power Project – Environmental Impact Review. Faculty of Natural Sciences, Comenius University, Bratislava, s. 241-245.
Šomšák, L., 2001: Optimalizácia vodného režimu ramennej sústavy z hľadiska prírodného prostredia –expertízne vyjadrenie. In Optimalizácia 2003.
Šomšák, L., Gazdík, M., Jankovičová, A., 1995: Dendrology of selected flood plain trees of the Danubian Lowland. In: Mucha, I. (ed.): Gabčíkovo Part of the Hydroelectric Power Project - Environmental Impact Review. Faculty of Natural Sciences, Comenius University, Bratislava, s. 155-160.
Šomšák, L., Šimonovič, V., Kollár, J., Lakatosová, E., 2002: Vypracovanie fytocenologickej mapy ľavostrannej inundácie Dunaja v úseku Dobrohošť – Sap. Sprievodná správa. Konzultačná skupina Podzemná voda, 37 s.
Šporka, F., 1980a: Informácia o makrozoobentose hlavných ramien Bačianskej ramennej sústavy v rokoch 1976, 1977. Zborník zo zasadnutia 22. zmiešanej komisie, Bratislava, s.85-89.
Šporka, F., 1980b: Výskyt a rozšírenie druhu Dreissena polymorpha Pallas vo vybranom úseku Bačianskeho ramena a zachytenie sprievodných druhov fauny dna. Záverečná správa subetapy. LRH Bratislava, 19 s.
Šporka, F., 1982: Makrozoobentos hlavných ramien Bačianskeho ramenného systému a vplyv hydrologického režimu na bentickú faunu ramien. Zborník ref. 6. konferencie ČSLS, Blansko, s.110-115.
Šporka, F., 1983: Chaetogaster diversisetosus sp. n. a new species of Naididae (Oligochaeta) from Czechoslovakia. Věstn. čs. Spol. zool., 47, s. 137-139.
Šporka, F., 1984:Výskyt a rozšírenie druhu Dreissena polymorpha Pallas (Mollusca, Bivalvia) v hlavných ramenách Bačianskeho ramenného systému. Zborník zo zasadnutia 26. zmiešanej komisie, Plovdiv, s.197-202.
Šporka, F., 1992: Máloštetinavce (Oligochaeta) Dunaja a vybraných vodných biotopov v jeho inundačnom území na Slovensku (Záverečná správa čiastkového projektu v rámci riešenia projektu "Biodiverzita v ČSFR"), 7 s.
Šporka, F., 1994: Klasifikácia pôvodných vôd inundačného územia Dunaja v slovensko-maďarskom úseku na základe fauny máloštetinavcov. Zborník X. Limnologickej konferencie, Stará Turá, s. 217-220.
Šporka, F., 1998: The typology of floodplain water bodies of the Middle Danube (Slovakia) on the basis of the superficial polychaete and oligochaete fauna. Hydrobiologia, 386, s. 55-62.
Šporka, F., Krno, I., 1995: Zmeny v bentickej faune hlavného toku a vôd ležiacich v inundácii po sprevádzkovaní VD Gabčíkovo. Zborník “Výsledky a skúsenosti z monitorovania bioty územia ovplyvneného vodným dielom Gabčíkovo”. Ústav Zoológie a ekosozológie SAV Bratislava, s.132-143.
Šporka, F., Nagy, Š.,1998: The macrozoobenthos of parapotamon-type side arms of the Danube river in Slovakia and its response to flowing conditions. Biológia, Bratislava, 53, s. 633-643.
Štepanovičová, O., 1989: Heteroptera v cenotických štruktúrach. Práce Slov. Ent. Spol. SAV, Bratislava, 7, s. 131 – 136.
Štepanovičová, O., 1991: Formation of taxocoenoses of Heteroptera in plant communities of relatively unaffected ecosystems and those affected by human activities, Acta F.R.N. Univ. Comen., Zoológia, 35, s. 31 – 51.
Štepanovičová, O., 1995a: Review of zoological research in the Danubian Lowland (terrestrial animals). In: Mucha, I. (ed.): Gabčíkovo part of the hydroelectric power project - environmental impact review. Faculty of Natural Sciences, Comenius University, Bratislava, Slovakia , s. 181-186.
Štepanovičová, O., Lapková, Z., 1984: Spoločenstvá bzdôch (Heteroptera) niektorých charakteristických biotopov záujmového územia Dunajského vodného diela. Práce Slov. ent. spol. SAV, Bratislava, 4, s. 189-203.
Tajovský, K., 1999: Impact of inundations on terrestrial arthropod assemblages in Southern Moravian floodplain forests, the Czech Republic. Ekológia (Bratislava), 18, Supplement 1, s. 177-184.
Takáč, J., 2001: Dôsledky zmeny klímy na bilanciu vody v poľnohospodárskej krajine. Národný klimatický program Slovenskej Republiky NKP 10/01, MŽP, SHMÚ, s. 16-26.
Tirjaková, E., 1992: Ciliophora of the riverside zone of the Danube river system. Acta Zool. Univ. Com., 36, s. 71–80.
Tirjaková, E., 1999: Nové nálezy cicaviek (Suctoria) na území Slovenska. Folia faunistica Slovaca, 4, s. 11-14.
Tkáčová H., Kováčik M., et al., 1996: Podunajsko DANREG, Záverečná správa, Ministerstvo životného prostredia SR.
Trpiš, M., 1957: Predbežný prehľad vážok (Odonata) Žitného ostrova. Biológia, Bratislava, 12, s. 433-447.
Trpiš, M., 1962: Ökologische Analyse des Stechmücken Population in der Donautiefebene in der Tschechoslowakei. Biologické práce, 8, s. 1-132.
Tuf, H., I., 2000: Communities of centipedes (Chilopoda) in three floodplain forests of various age in Litovelské Pomoraví (Czech Republic). Fragmenta faunistica (Warszawa), 43 (Supplement), s. 327-332.
Uherčíková, E., 1995: Fytocenologické a ekologické pomery lesov inundácie Dunaja. Kand. diz. práca, Ms. depon. in PriFUK a ÚZ SAV, Bratislava, 221 s.
Uherčíková, E., 1998: Transformation changes in the association Salici-Populetum in the inundation area of the Danube river (Slovakia). - Biologia (Bratislava), 53, s. 53-63.
Uherčíková, E., 2001: Expertízne vyjadrenie k optimalizácii vodného režimu v inundácii Dunaja. Časť: Lesné fytocenózy v inundácii. In Optimalizácia 2003.
Úrad predsedu vlády MR, 1999: Kancelária vládneho komisára pre Dunaj: „Úlohy analýzy vplyvov pre Dunaj, odovzdaná SR v r. 1999.
Varga, L., 1993: Záchrana a reprodukcia listnatých drevín v meniacich sa hydrologických podmienkach. LVÚ Zvolen, rukopis.
Vaškovská, E., 1986: Litogeochemická charakteristika fluviálnych sedimentov a pôd holocénu v Podunajskej rovine. Regionálna geológia Západných Karpát, Správy o výskumoch GÚDŠ 21, Bratislava, s. 127-136.
Vaškovský, I., et al., 1982: Vysvetlivky ku geologickej mape juhovýchodnej časti Podunajskej nížiny 1:50000. GÚDŠ Bratislava, 115 s.
Vojtuš, M., 1986: Príspevok k riešeniu niektorých problémov pestovania topoľov na Slovensku. VÚLH Banská Štiavnica, VS Gabčíkovo. Referát.
Vranovský, M., 1974: Zooplanktón Bačianskeho systému ramien pred vyústením do hlavného toku a jeho význam pre formovanie zooplanktónu v Dunaji. Biologické práce (Bratislava), 20 (7), s. 1-80.
Vranovský, M., 1985: Zooplanktón dvoch hlavných ramien Bačianskej ramennej sústavy. (Dunaj, r. km 1820,5-1825,5). Práce Lab. Rybár. Hydrobiol. (Bratislava), 5, s. 47-100.
Vranovský, M., Illiová, M., 1999: Zooplankton in the Danube and its left side arm system. In: I. Mucha (ed): Gabčíkovo part of the hydroelectric power project - Environmental impact review. Faculty of Natural Sciences, Comenius University, Bratislava, s. 167-174.
Zulka, K., P., 1999: Terrestrische Arthropoden, p. 259-271; Anhang 6 - Bemerkenswerte Arthropoden der March-Thaya-Auen. In: Kelemen, J., Oberleitner, I. (Red.), Fließende Grenzen. Lebensraum March-Thaya-Auen. Umweltbundesamt, Wien, s. 377-381.
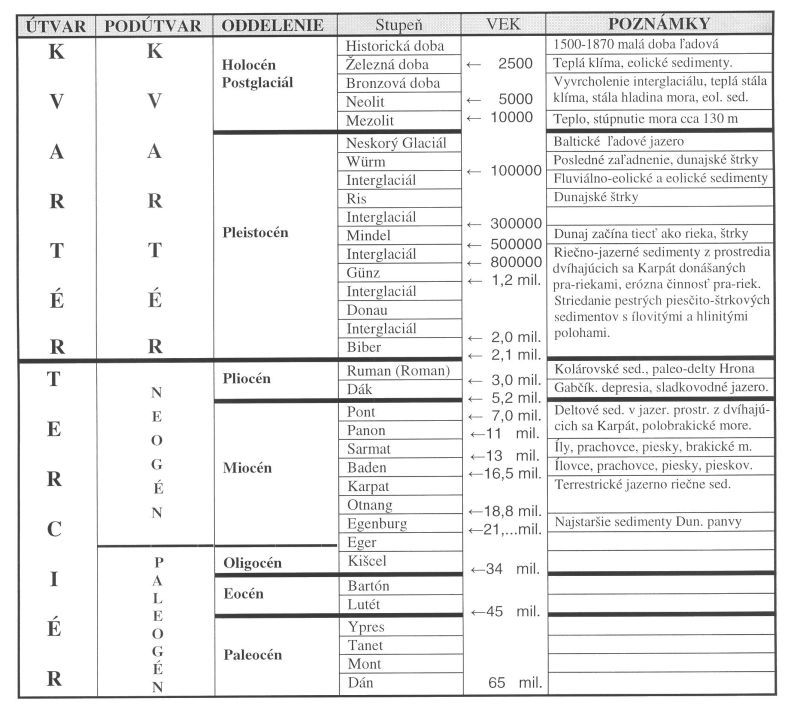
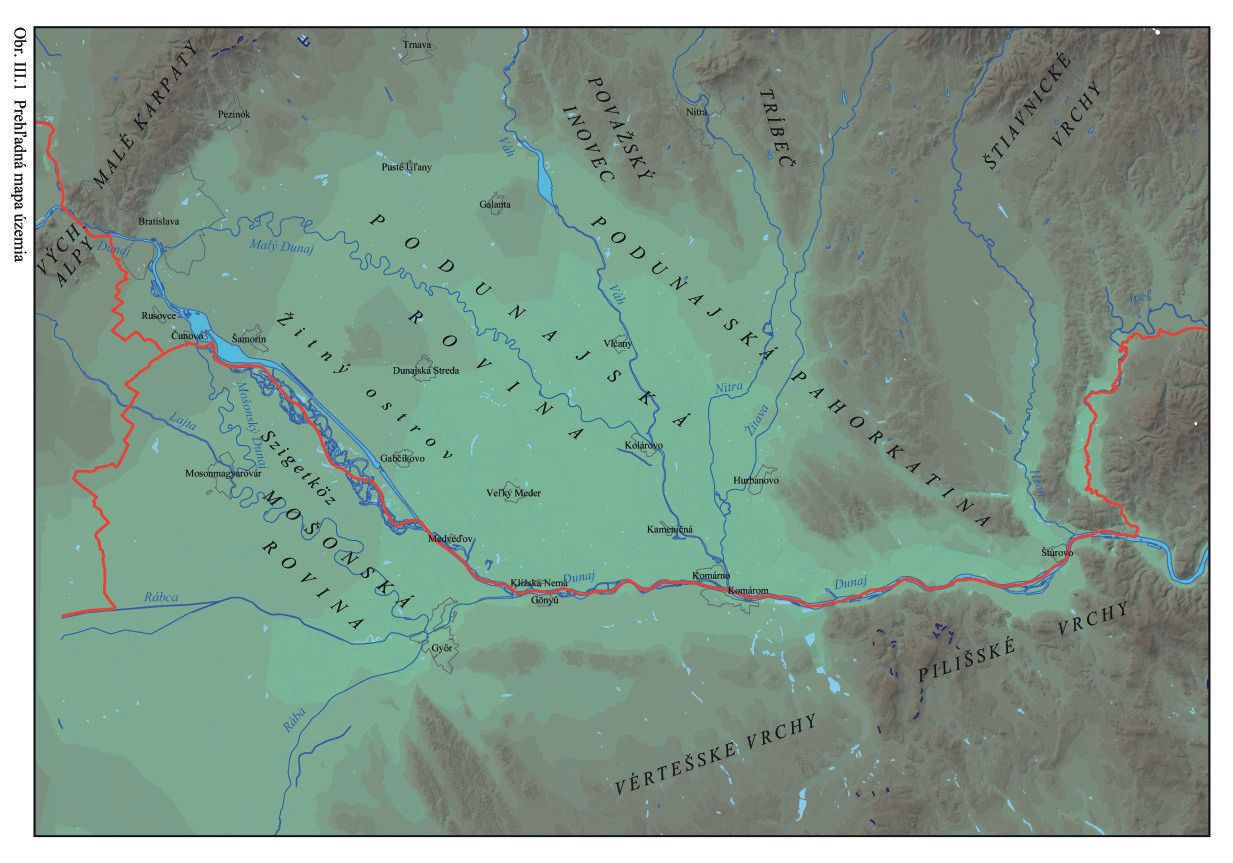{kind=link}
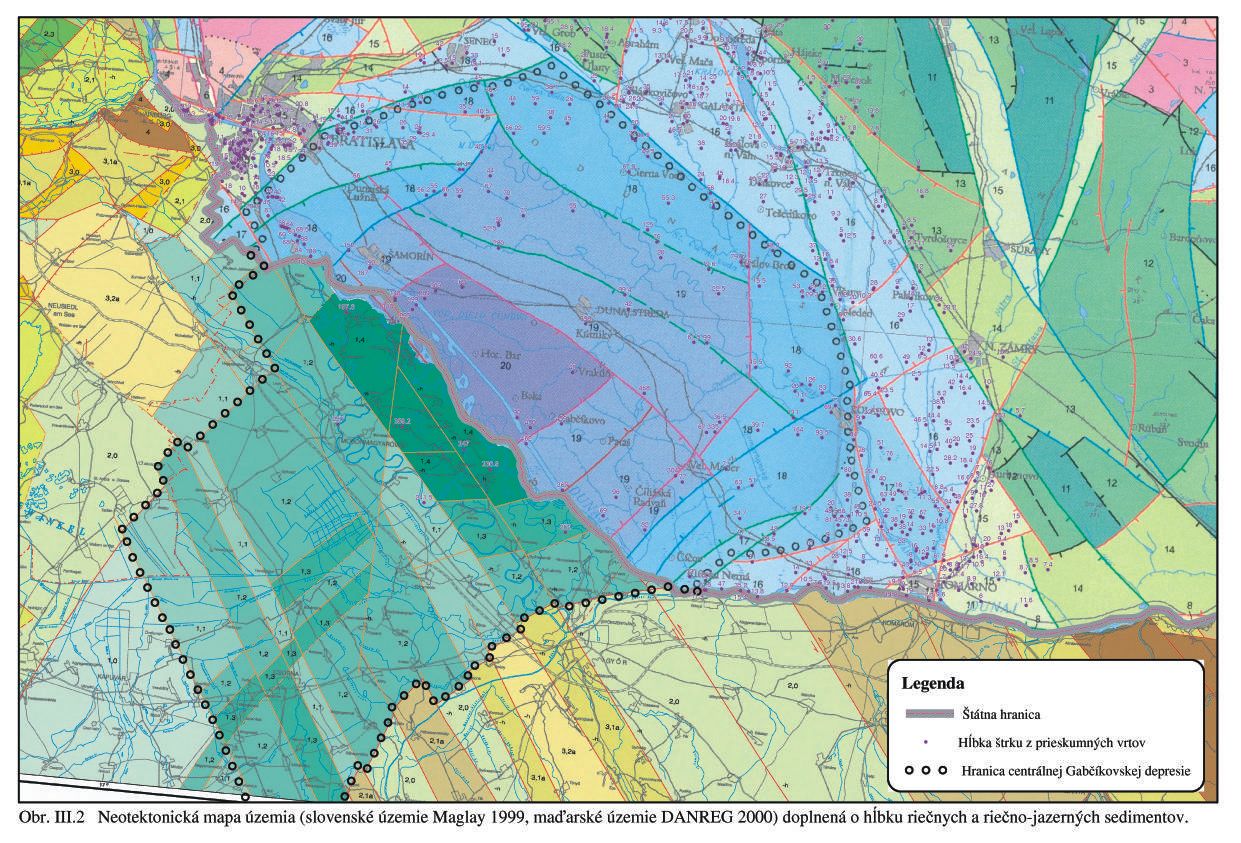{kind=link}
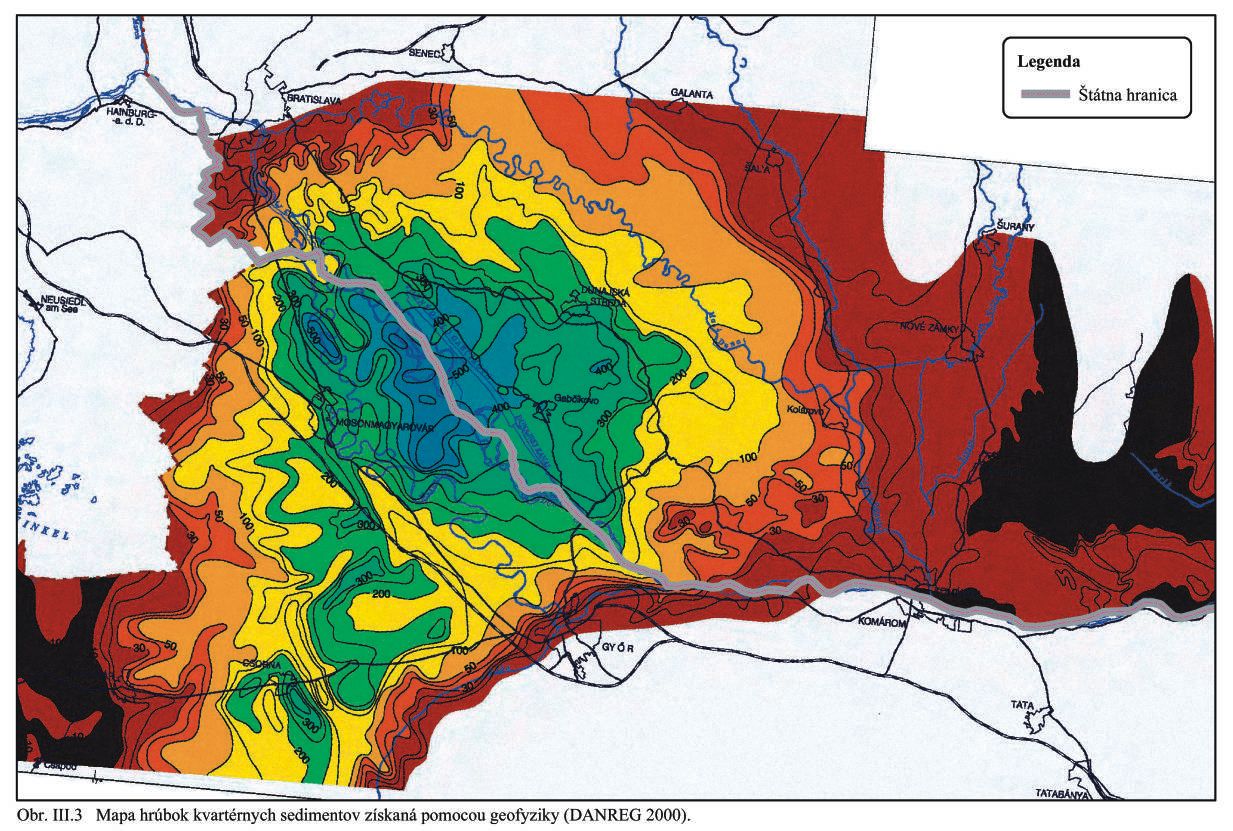{kind=link}
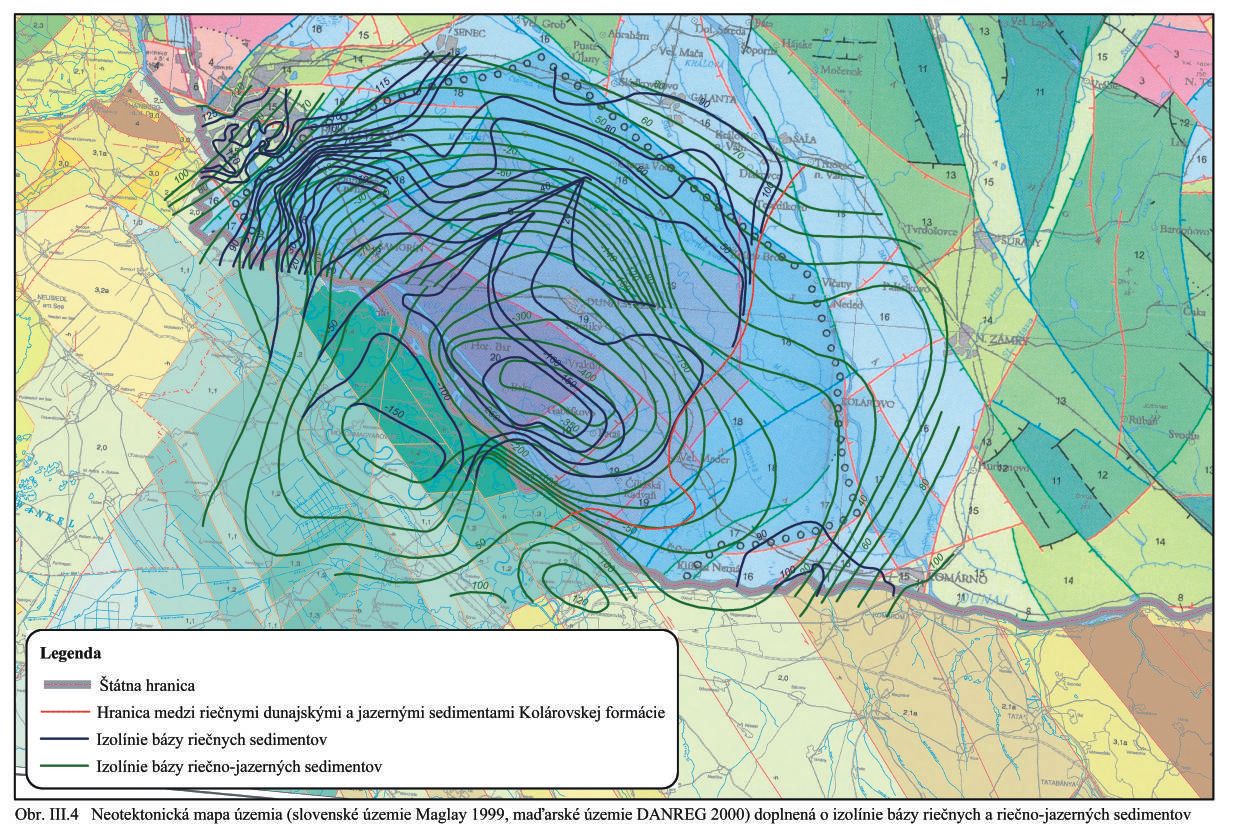{kind=link}
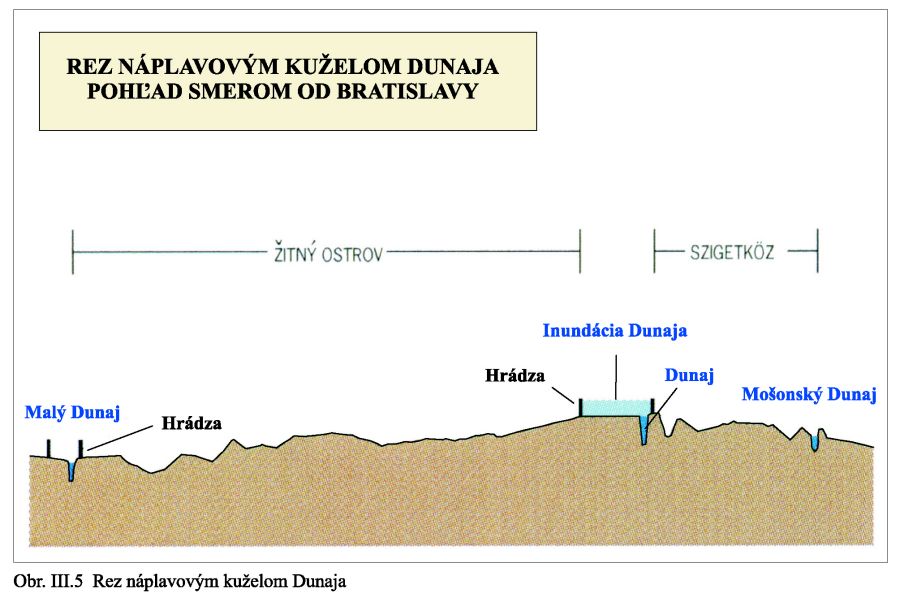{kind=link}
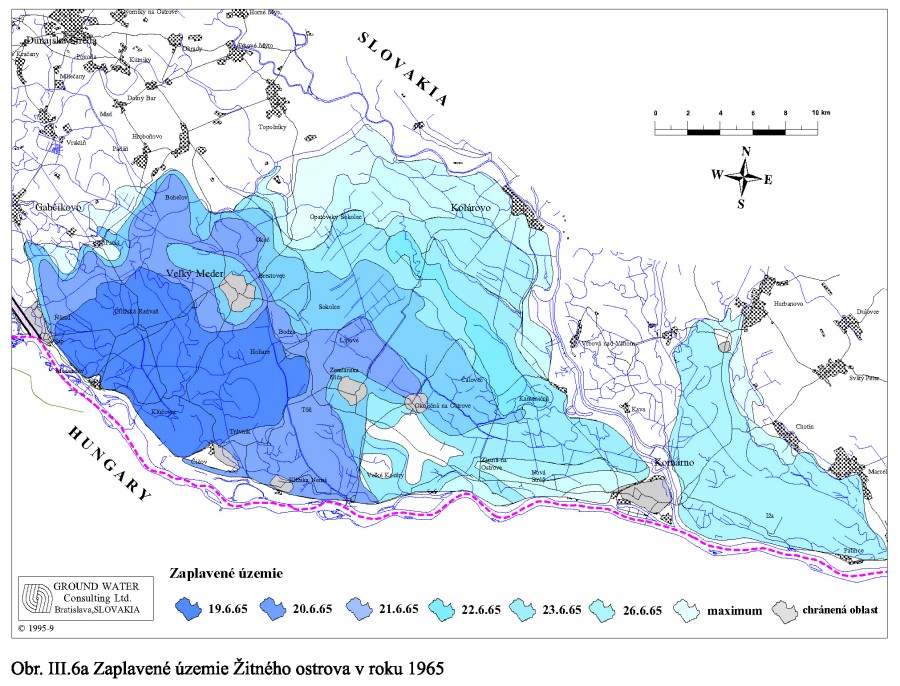{kind=link}
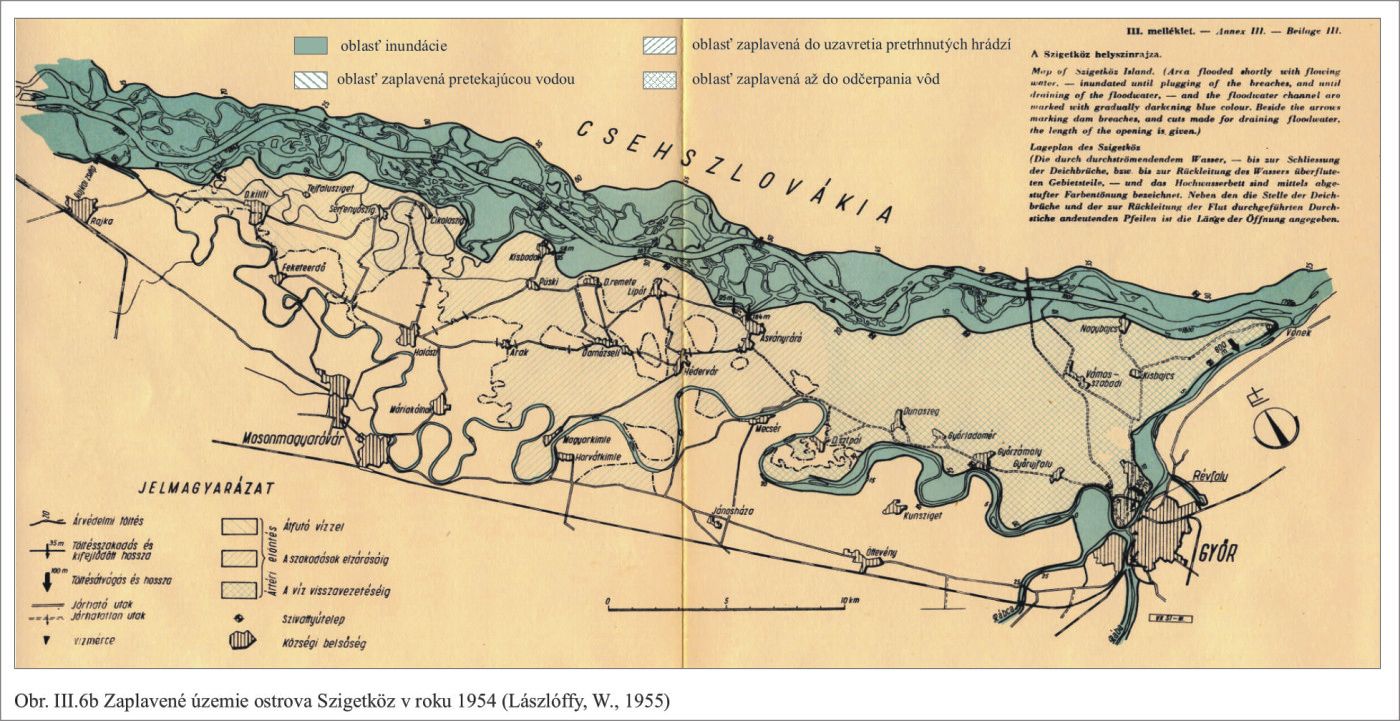{kind=link}
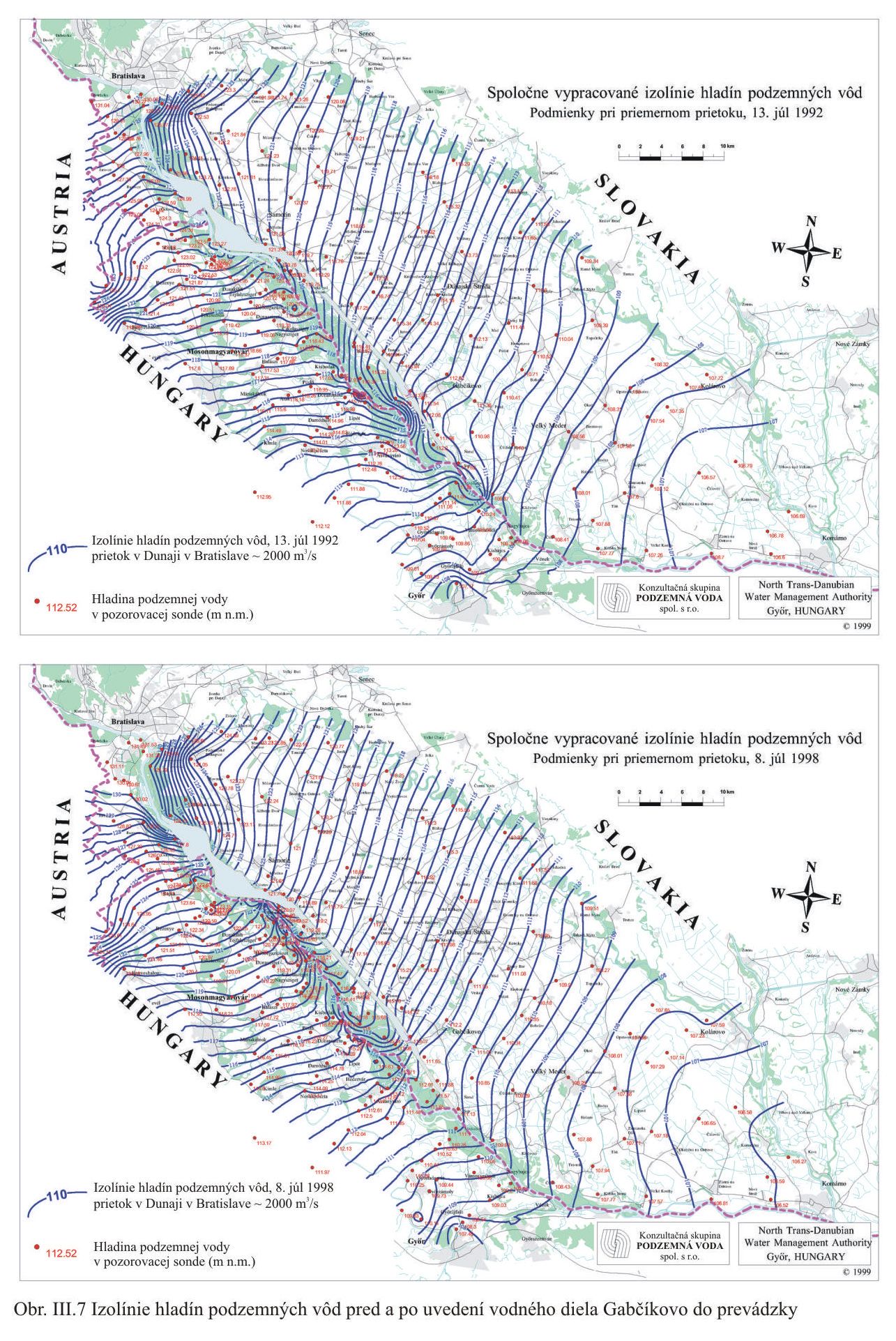{kind=link}
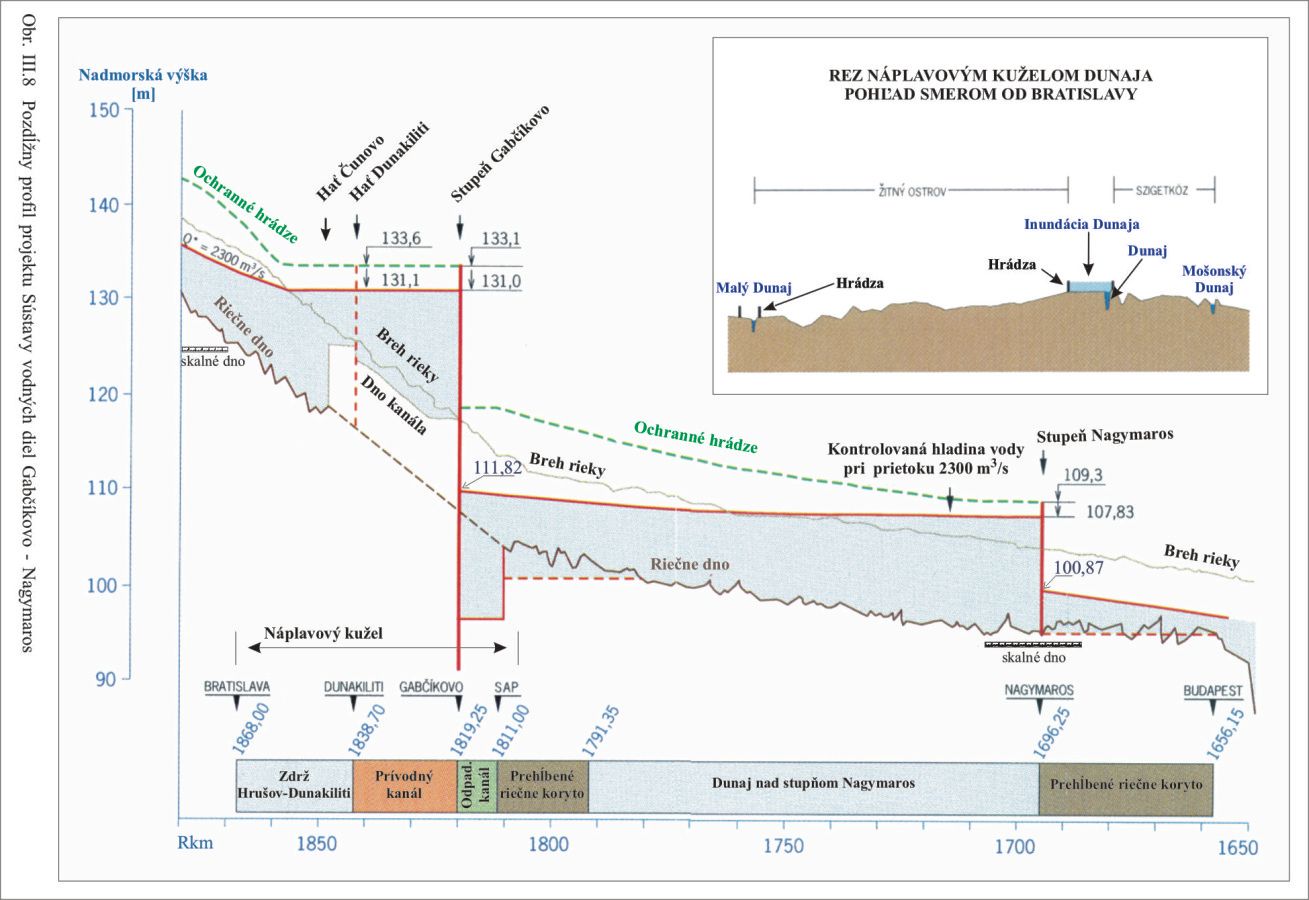{kind=link}
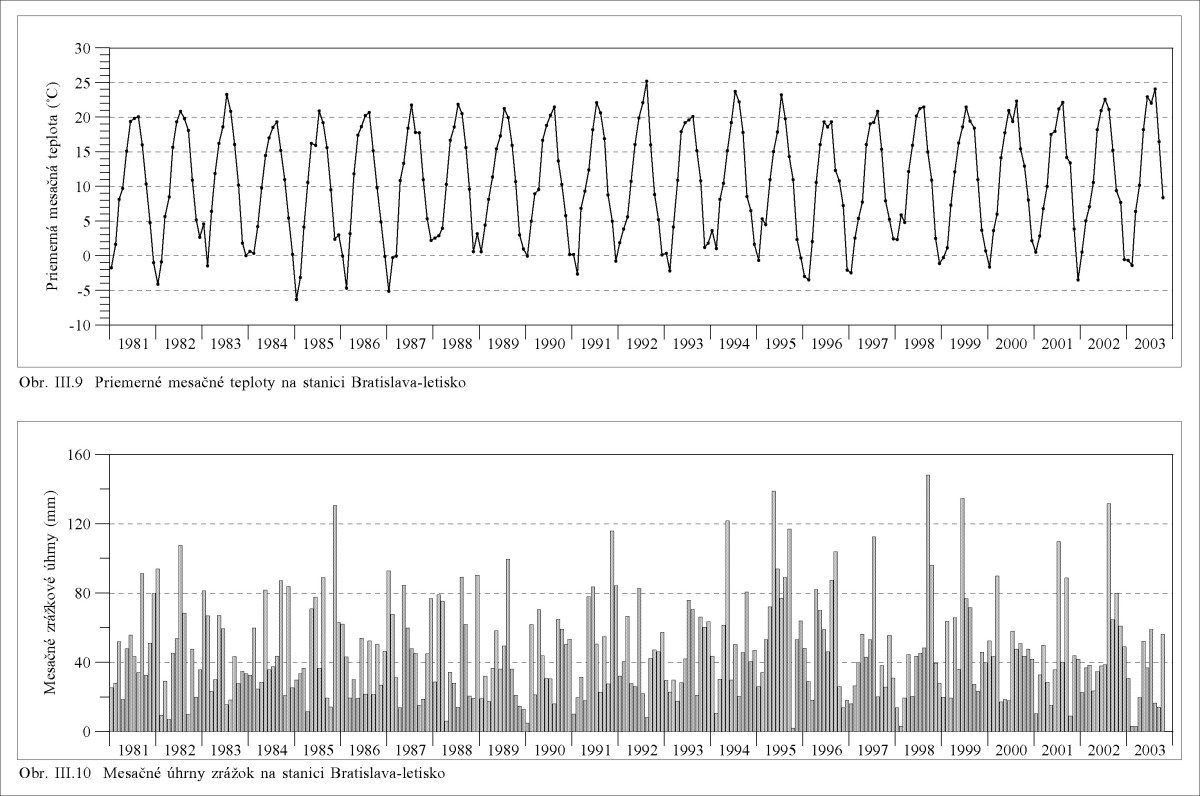{kind=link}
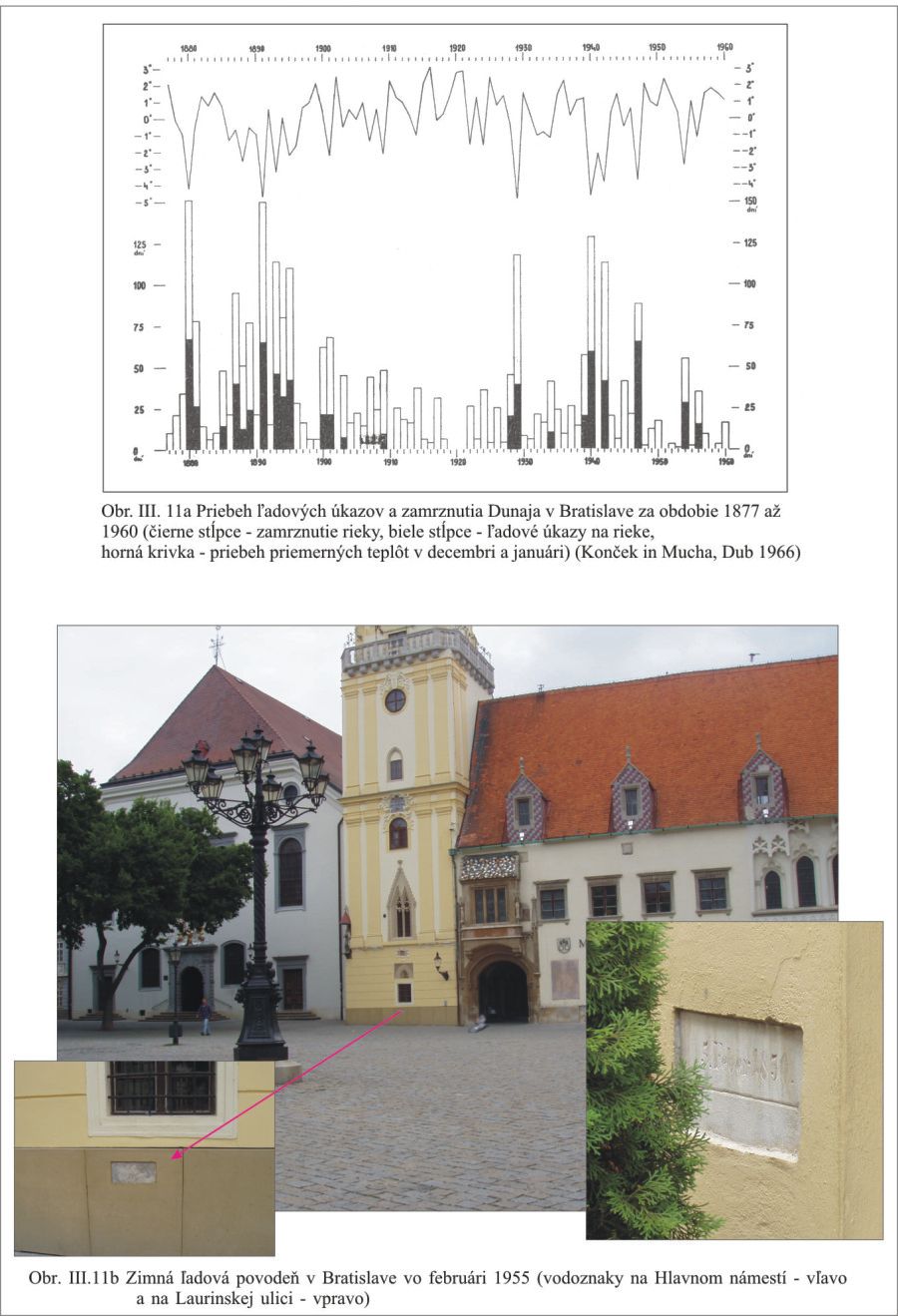{kind=link}
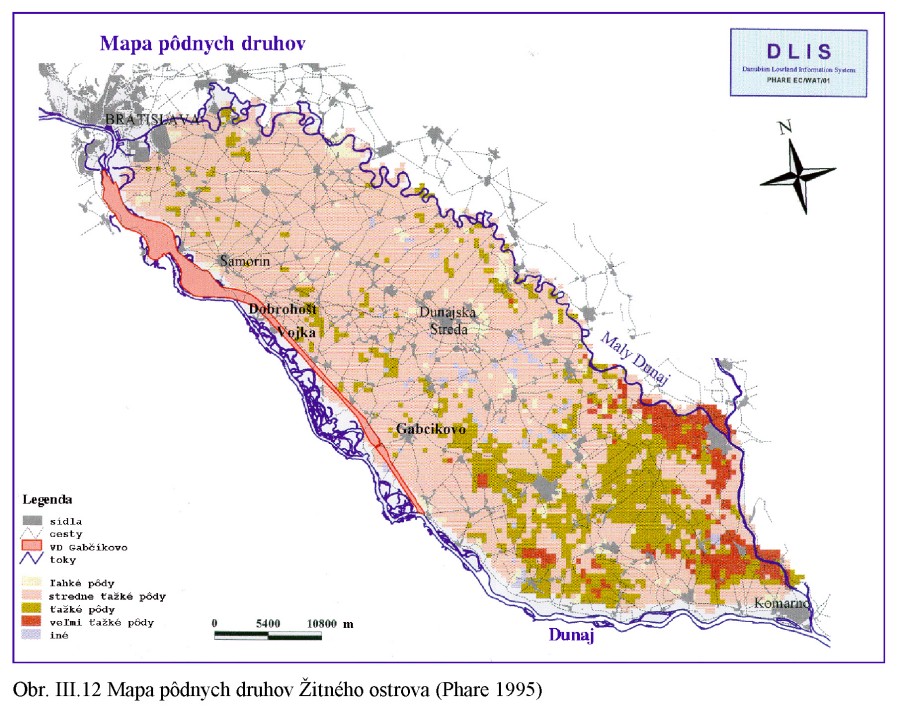{kind=link}
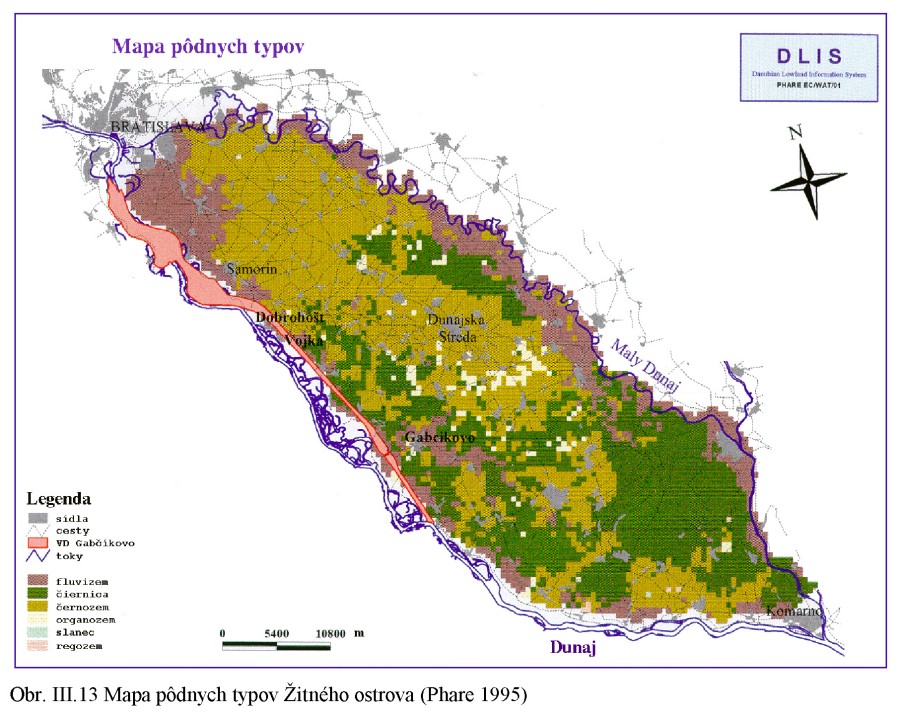{kind=link}
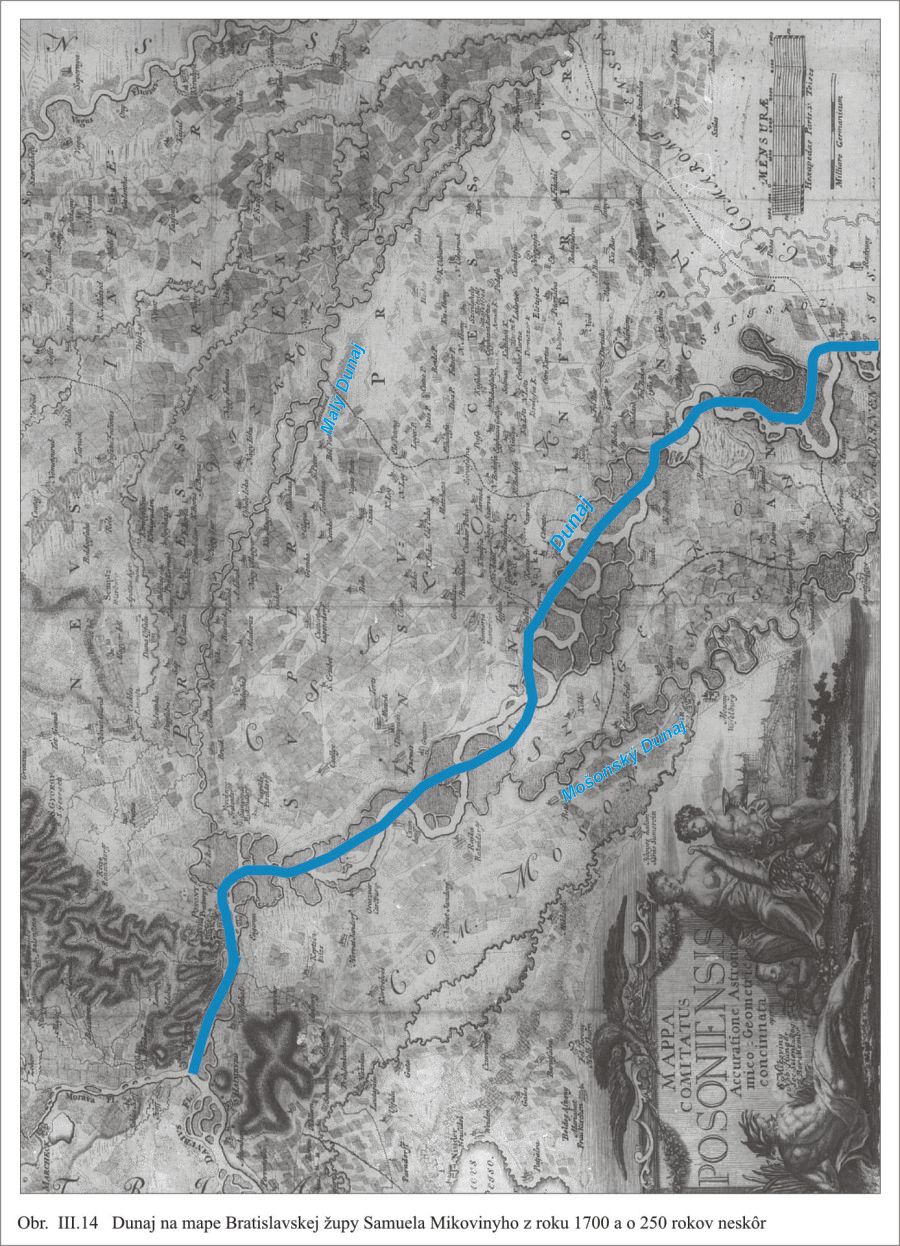{kind=link}
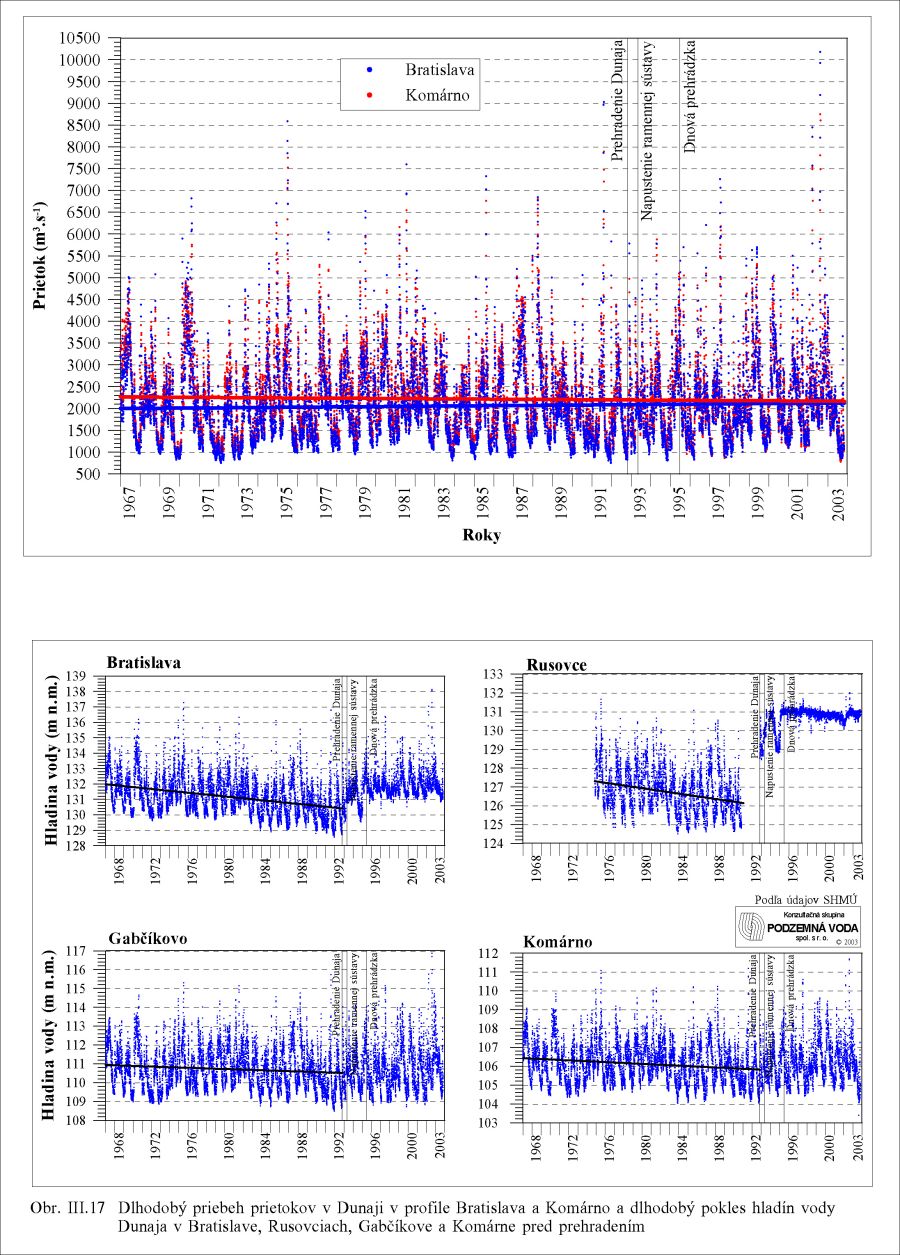{kind=link}
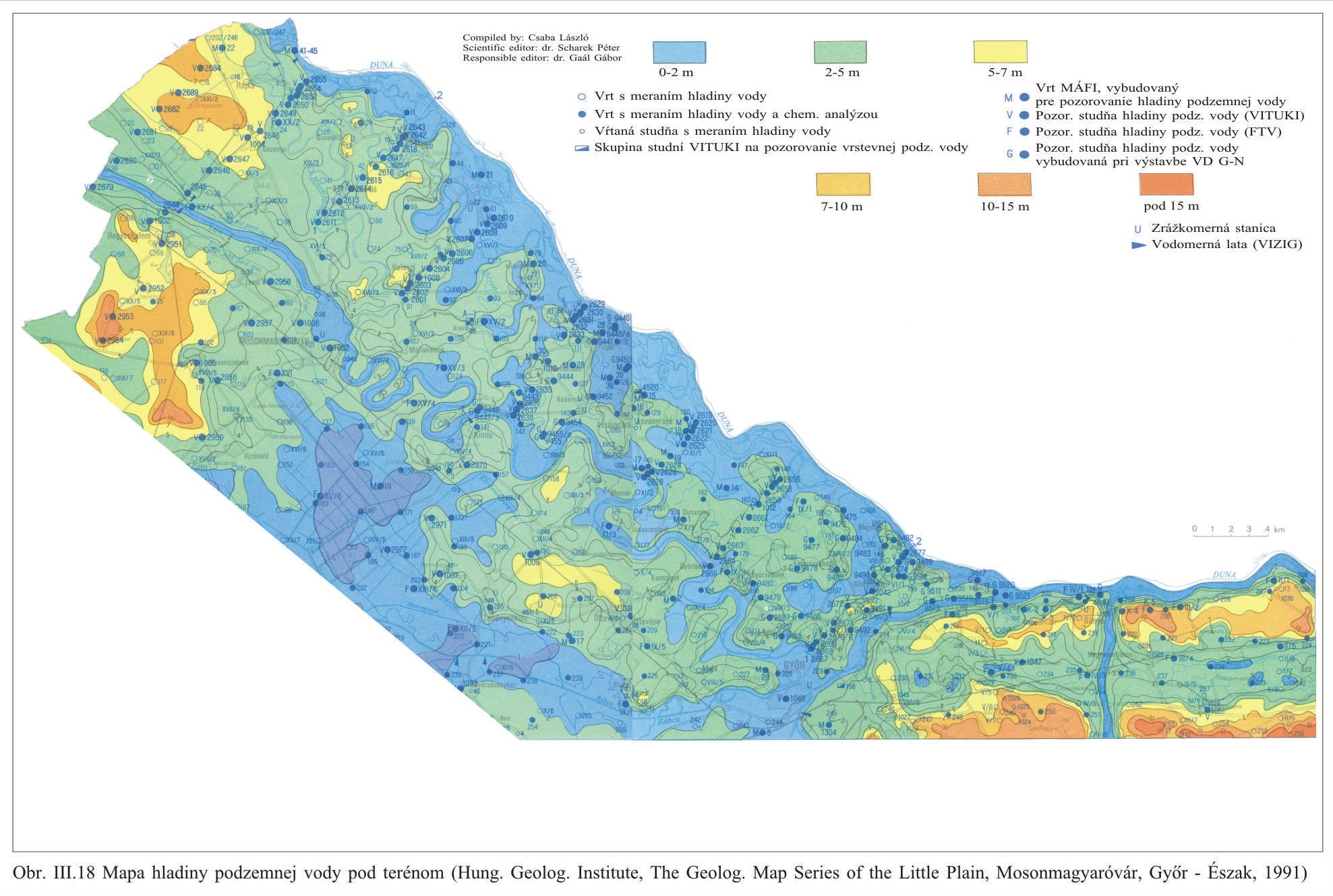{kind=link}
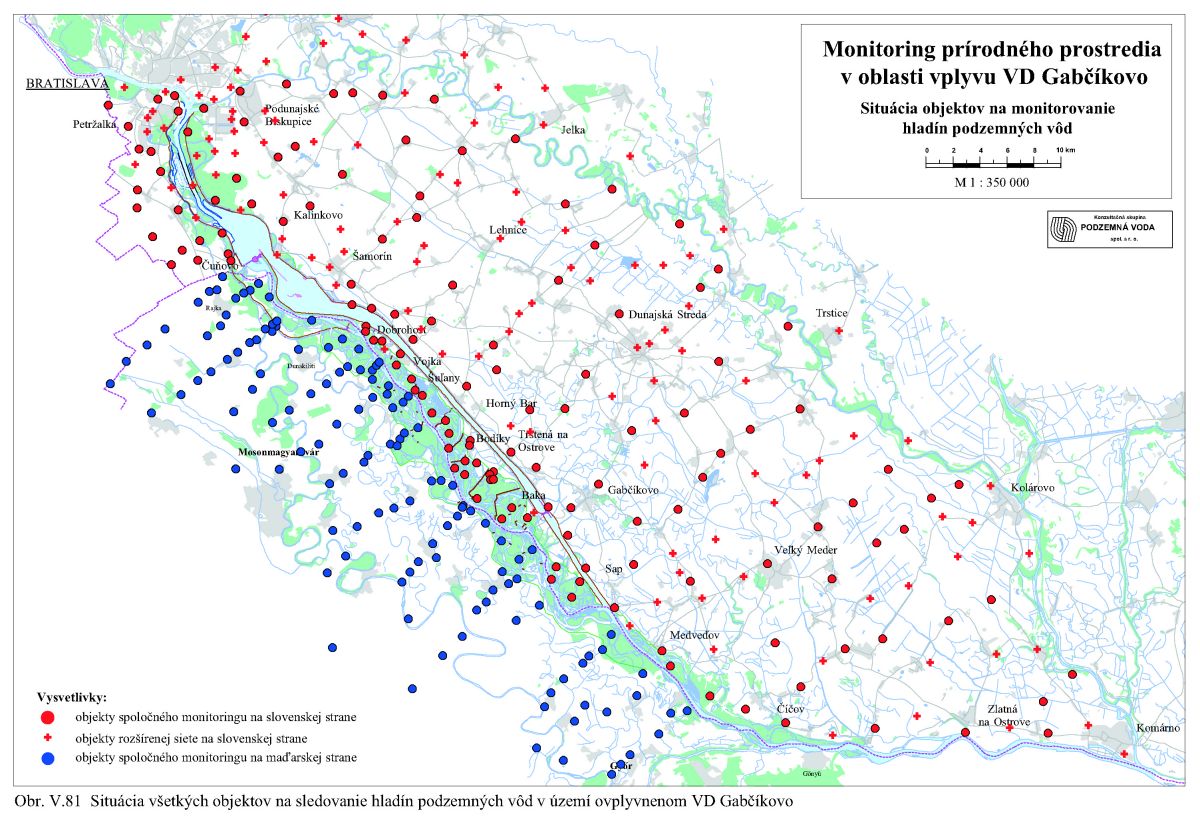{kind=link}
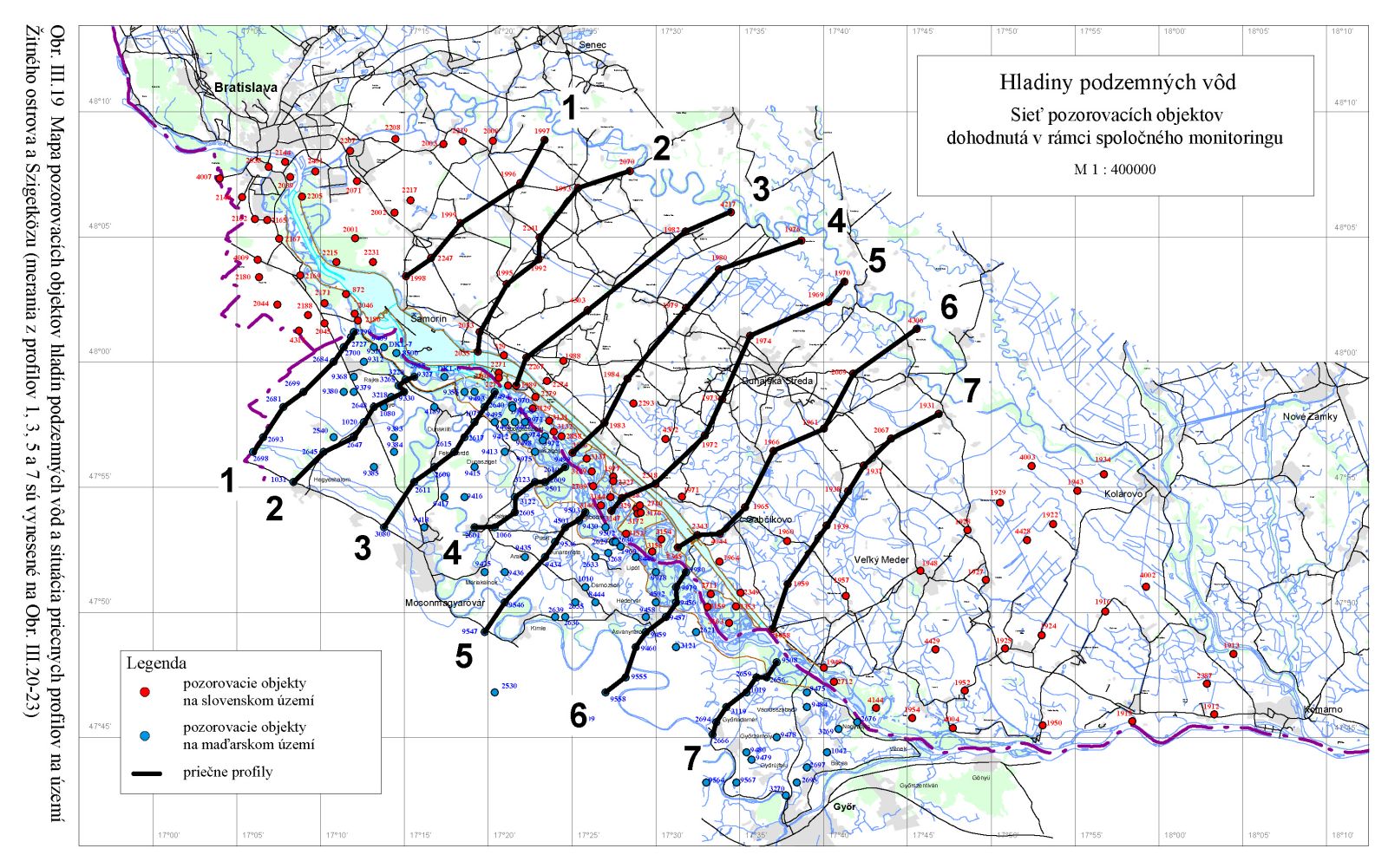{kind=link}
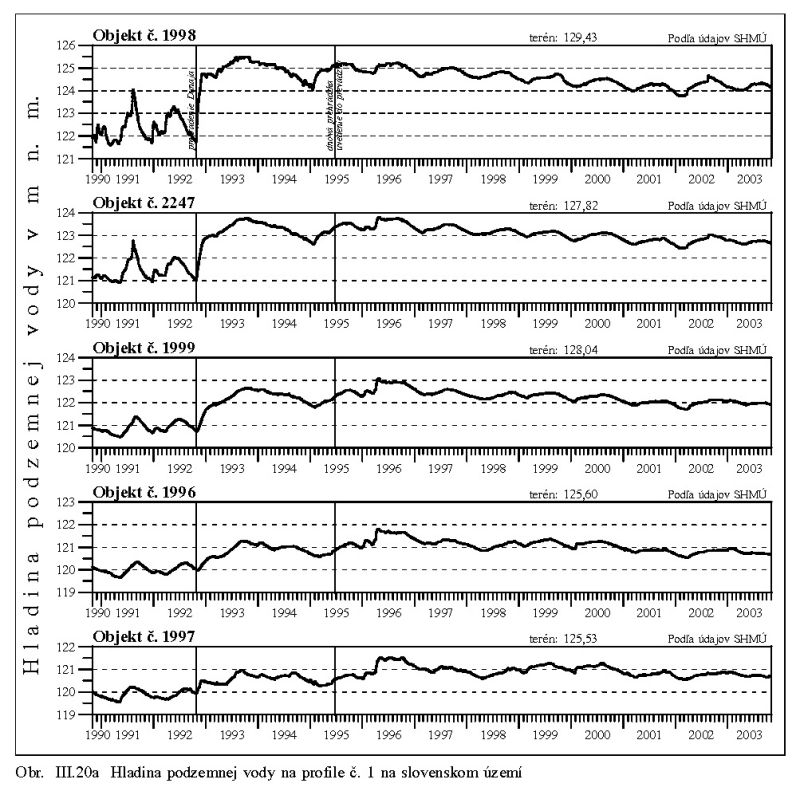{kind=link}
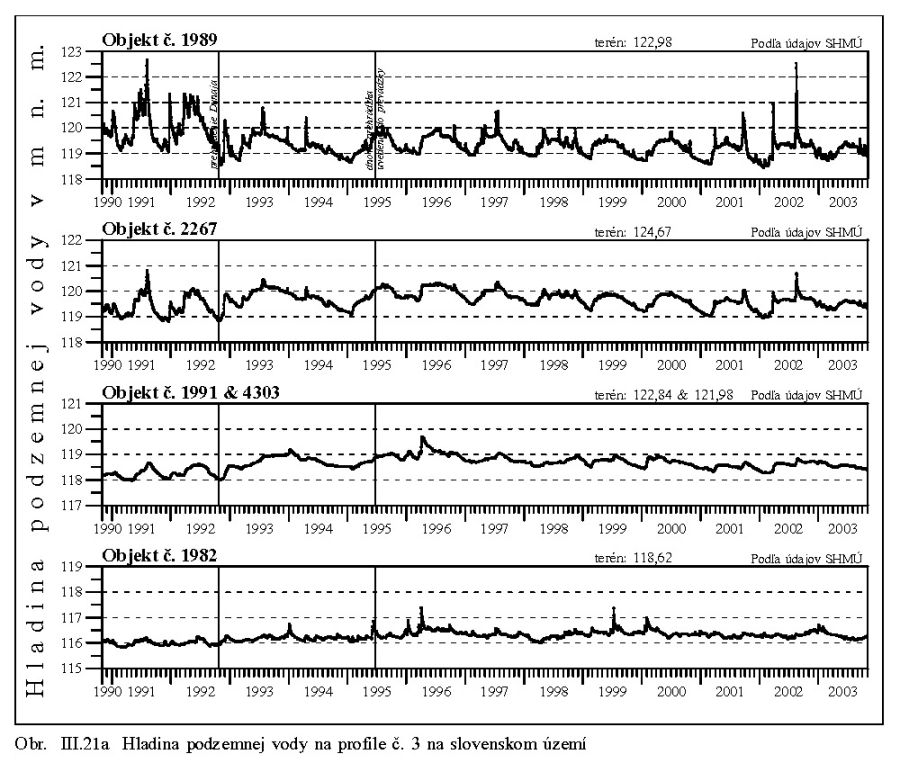{kind=link}
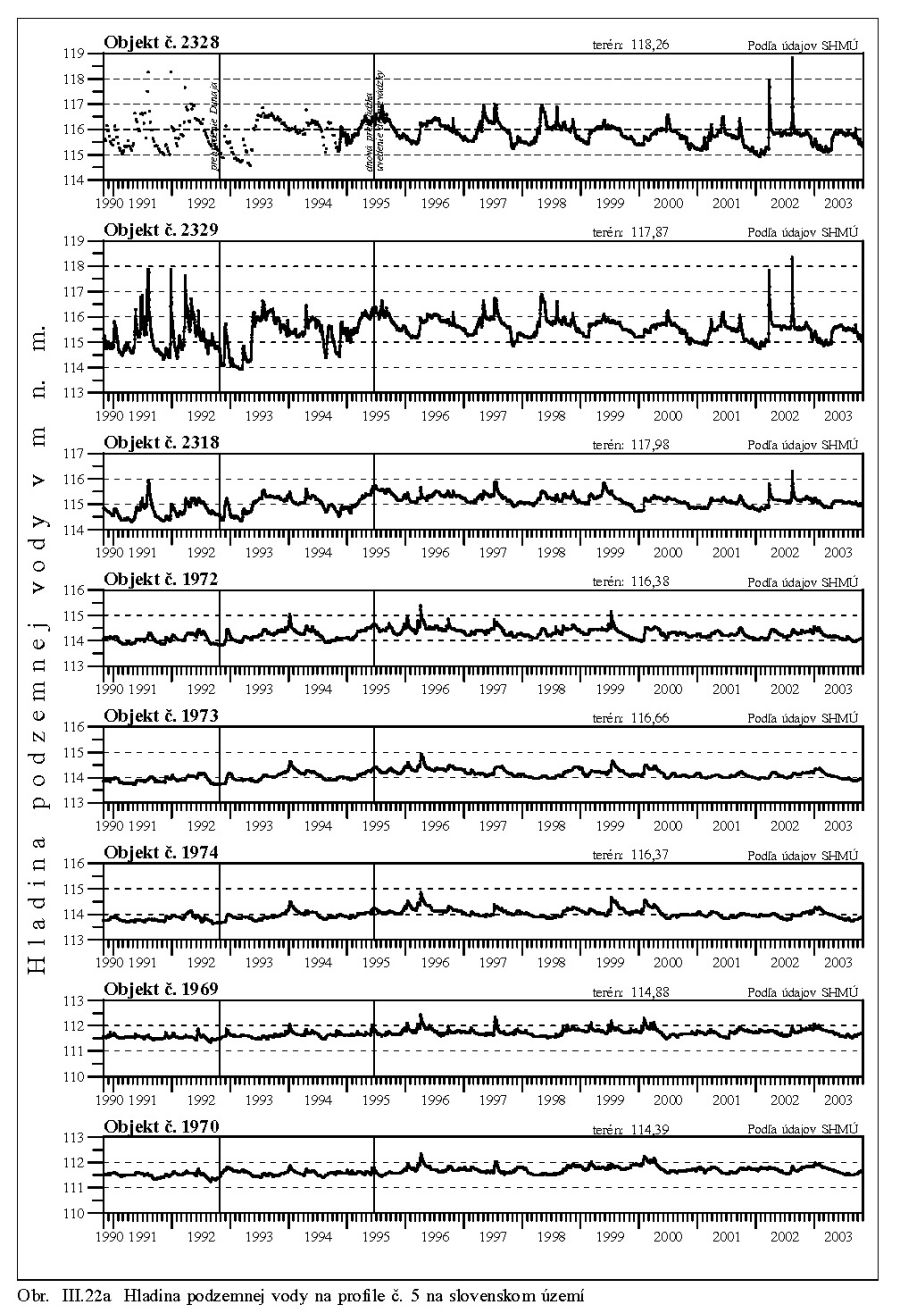{kind=link}
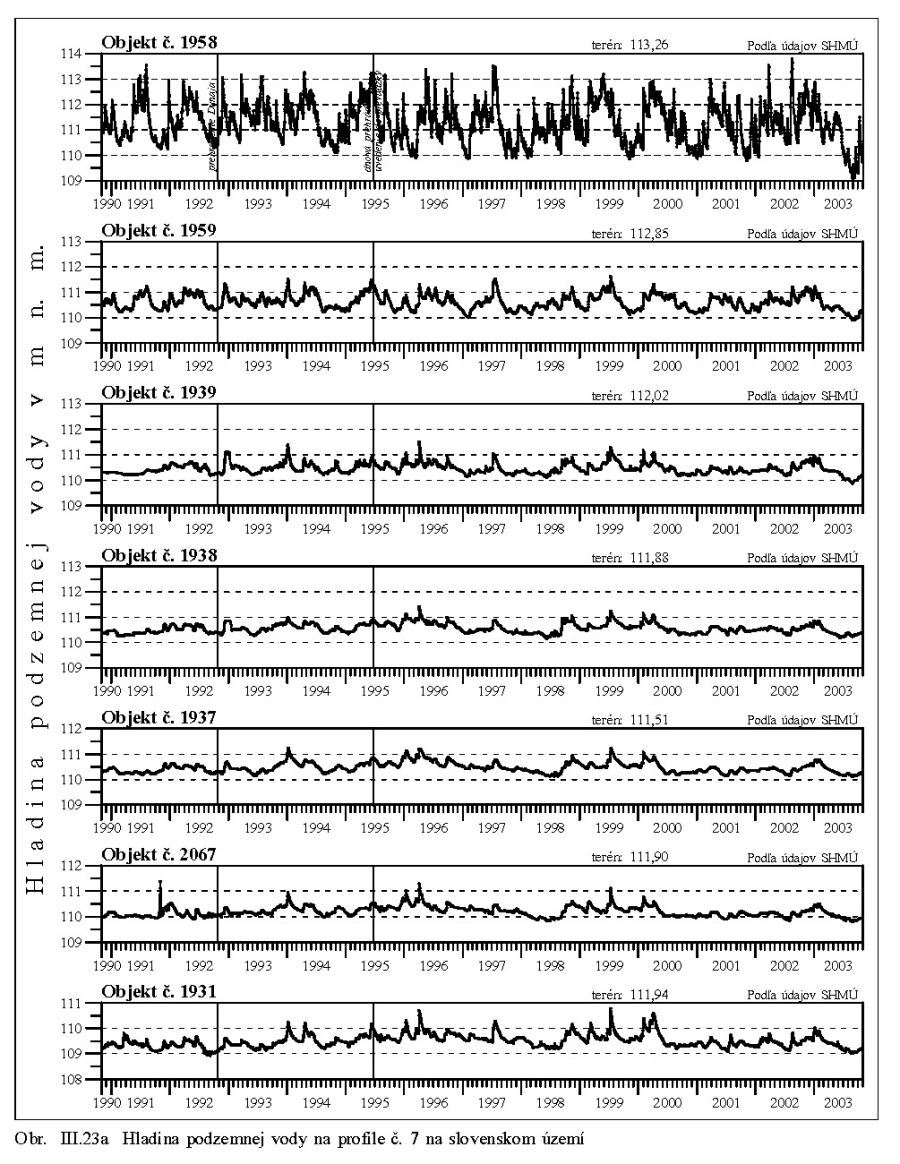{kind=link}
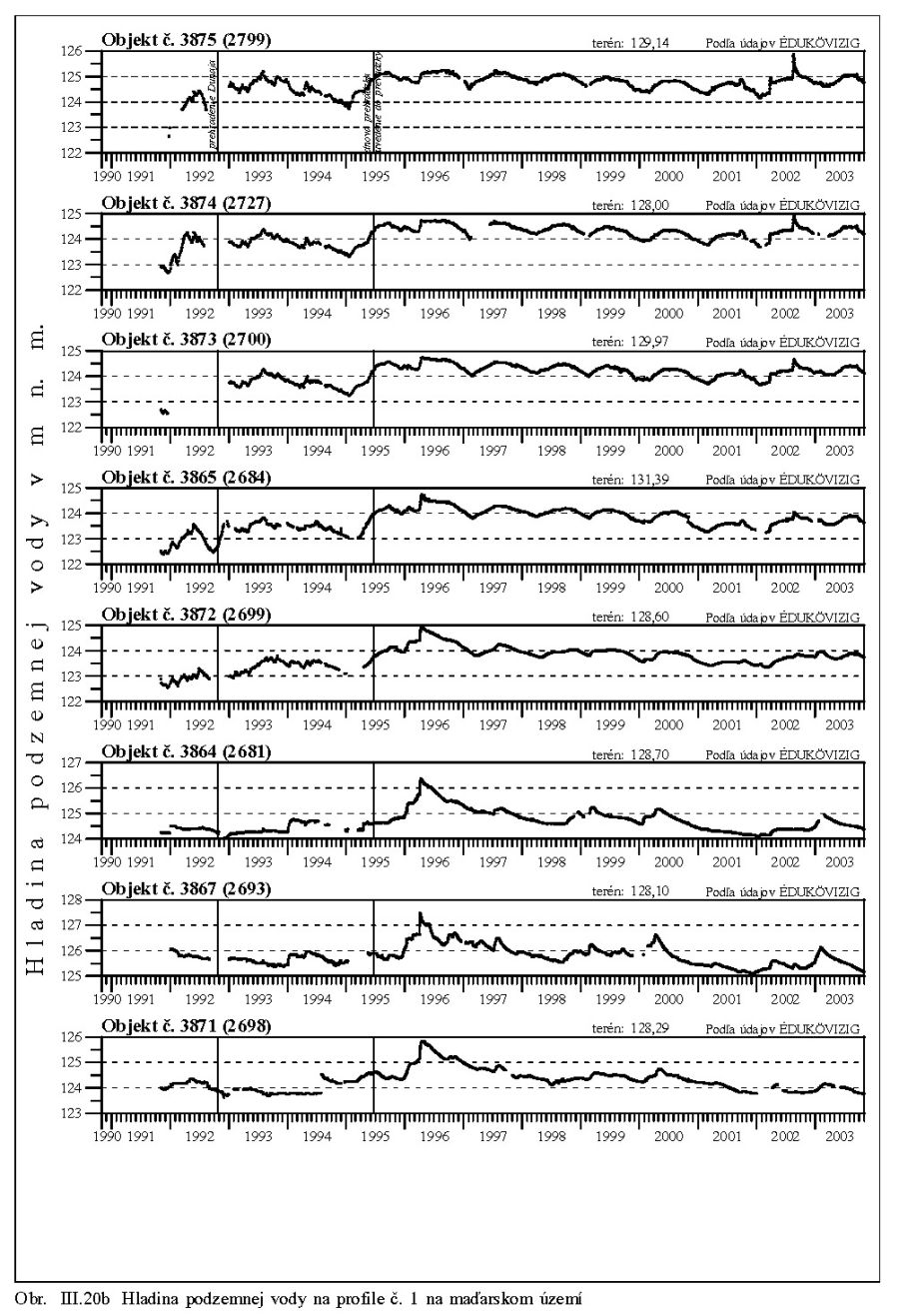{kind=link}
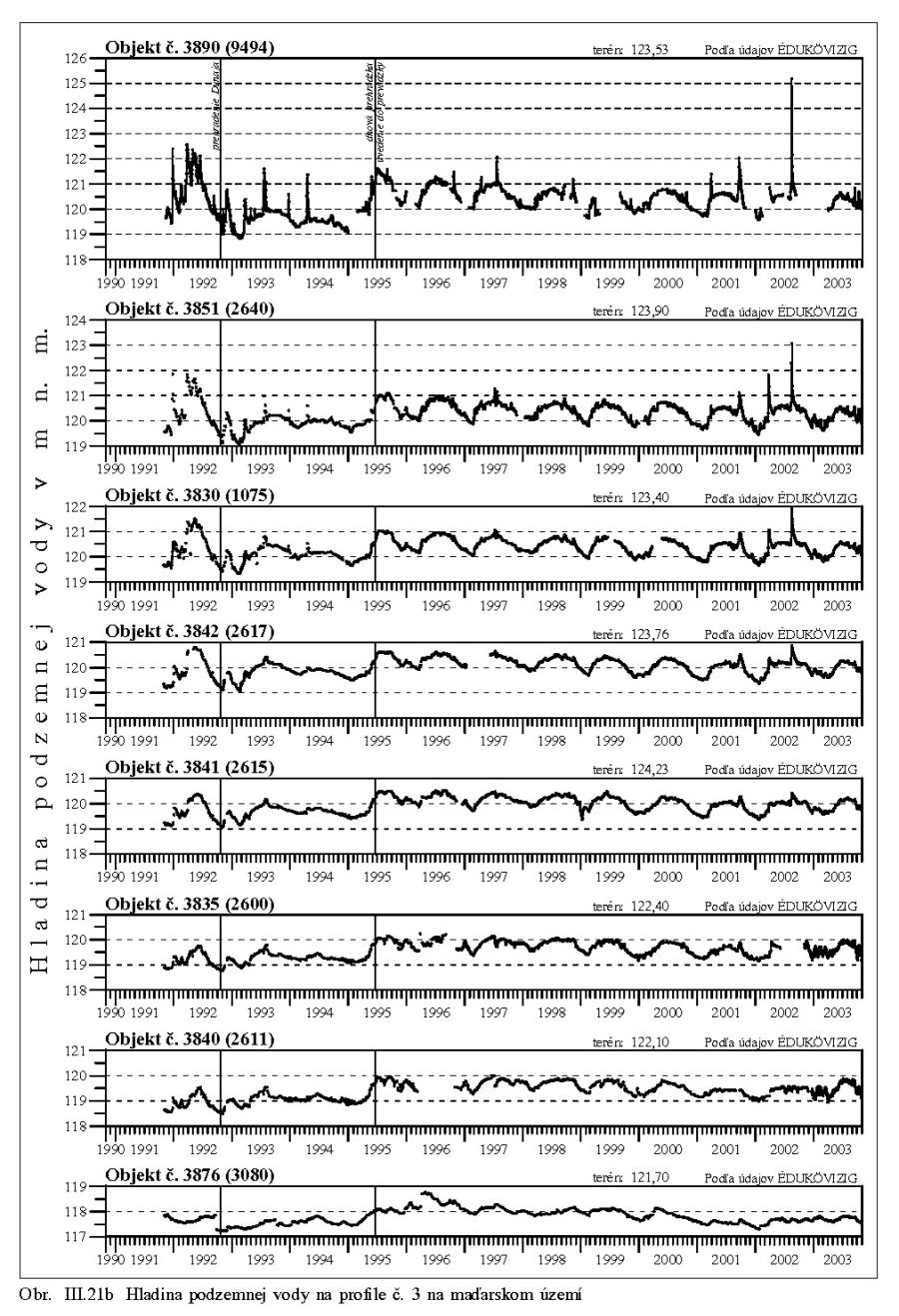{kind=link}
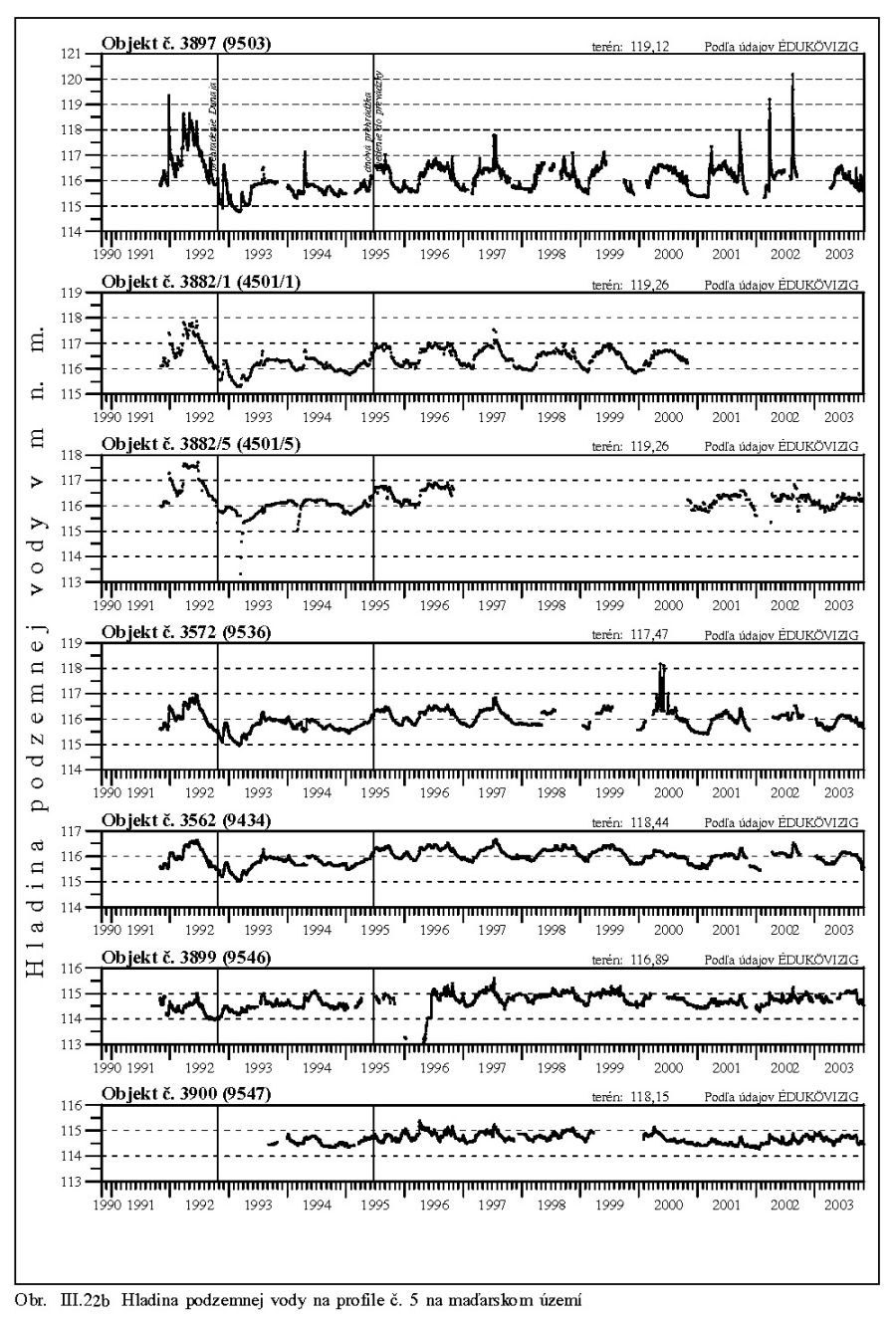{kind=link}
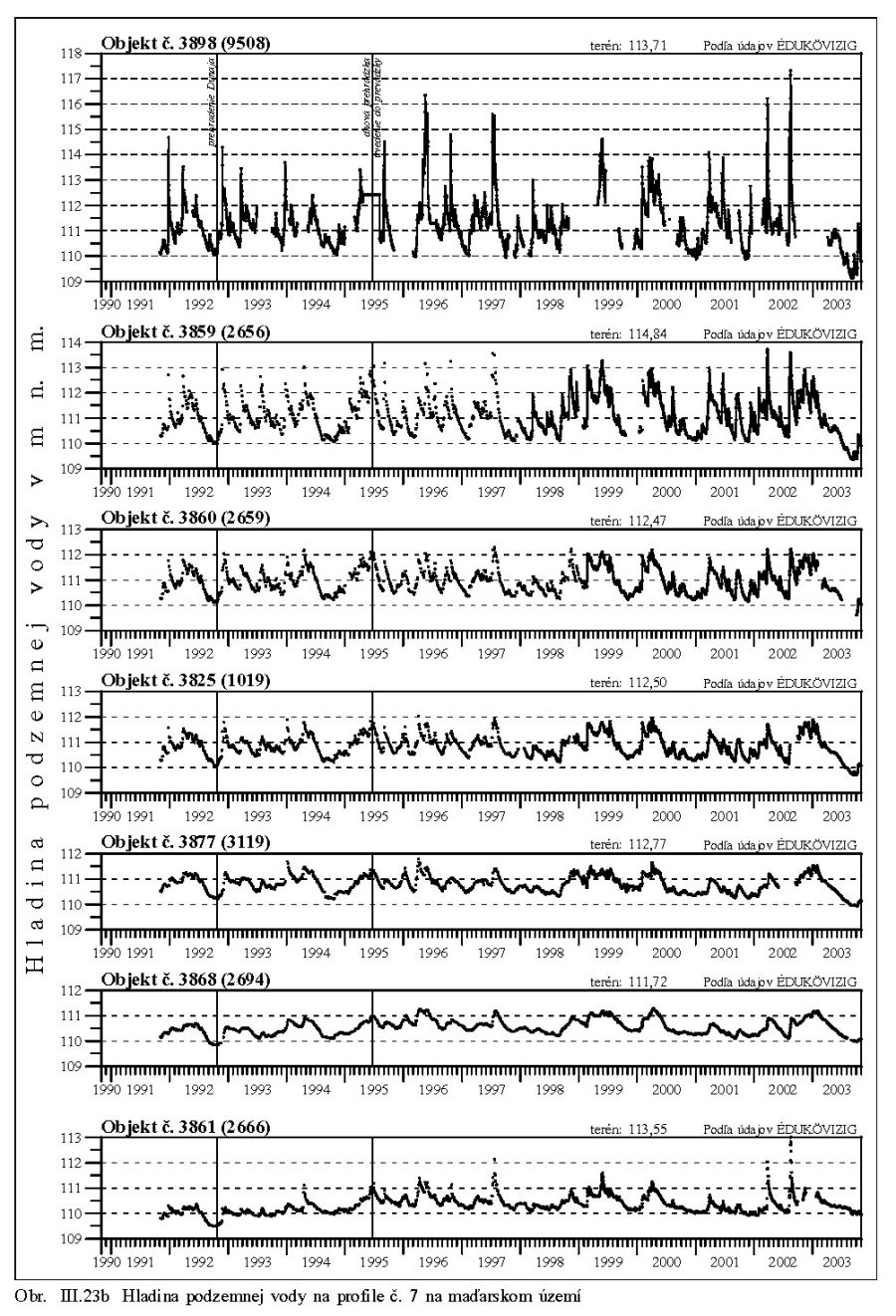{kind=link}
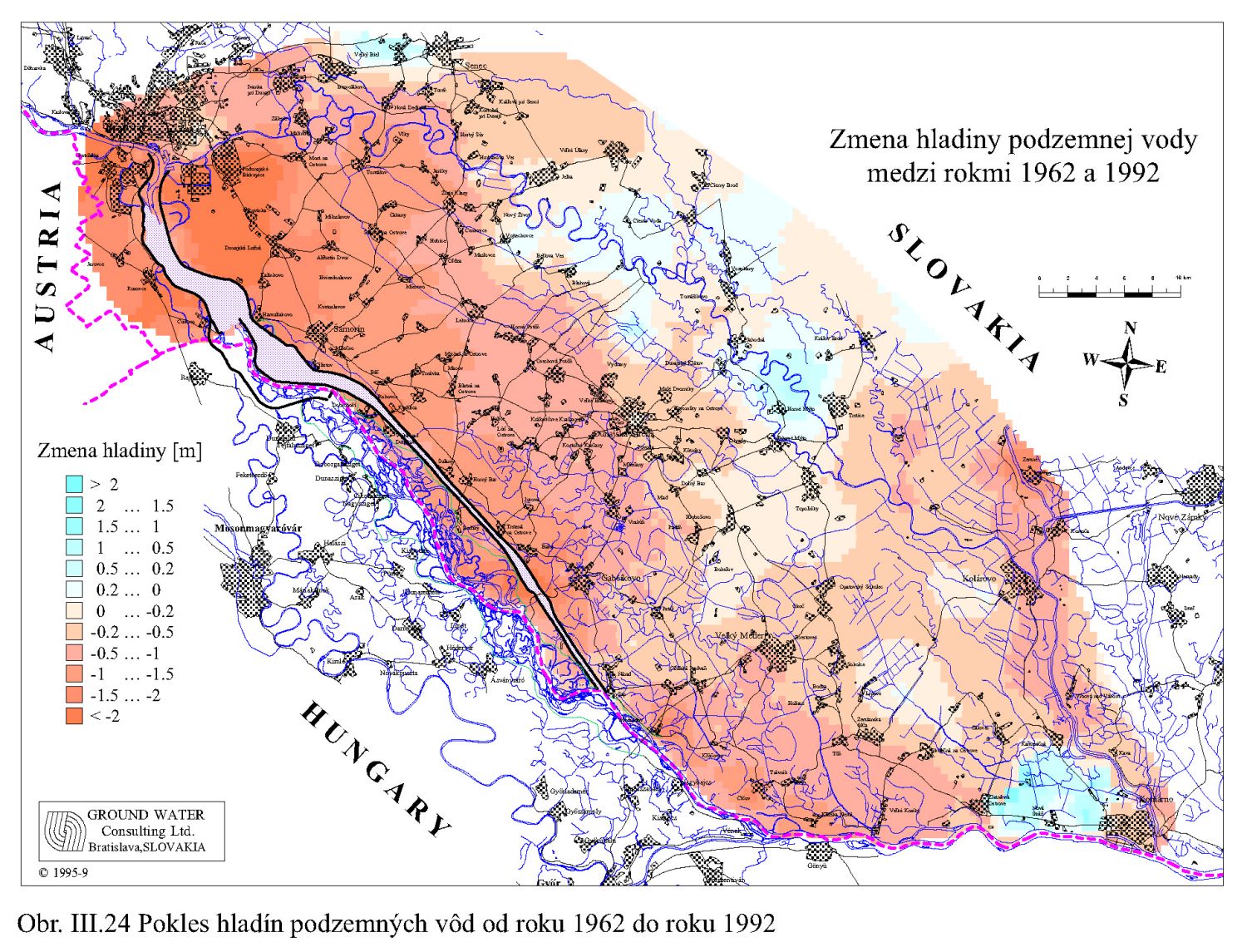{kind=link}
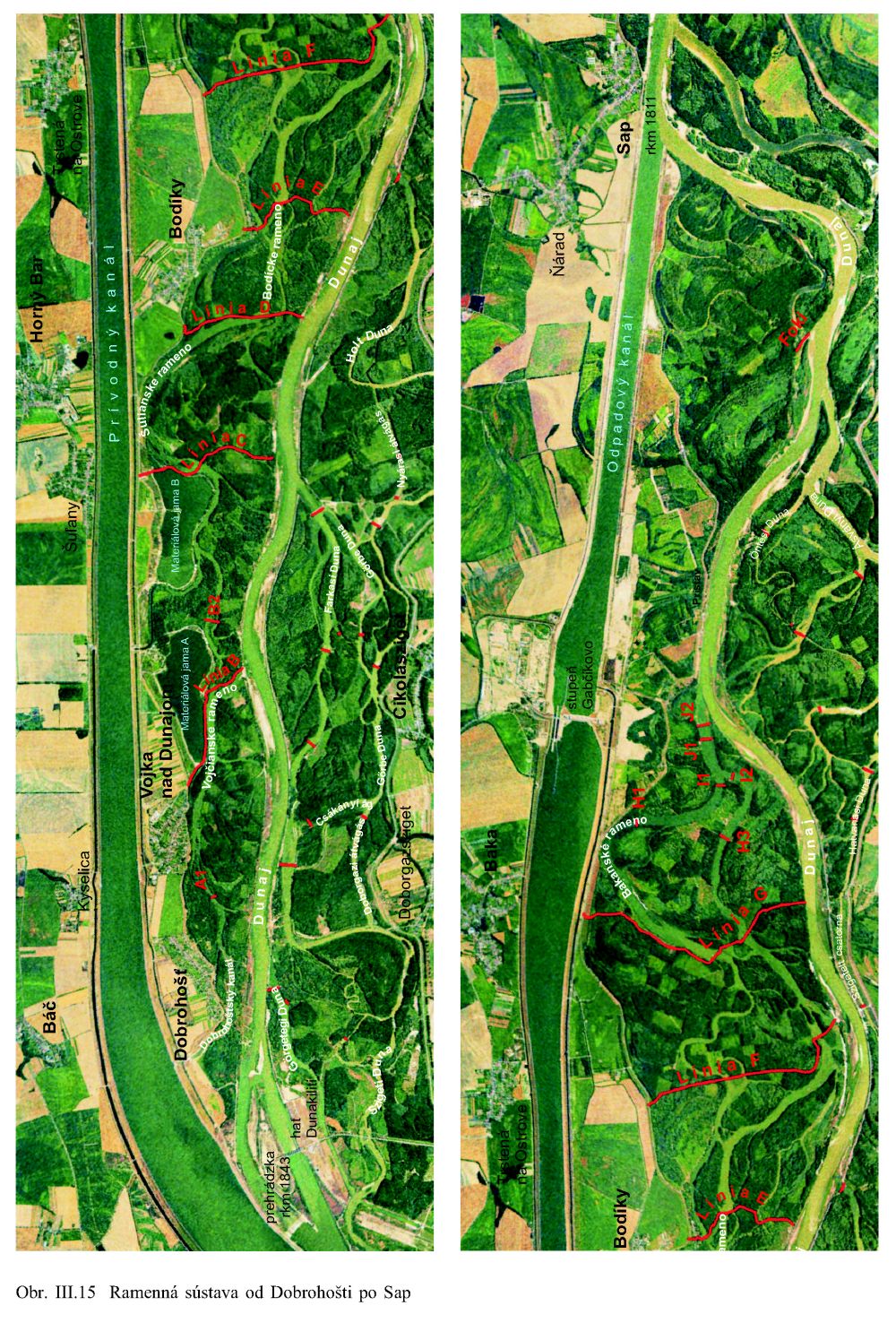{kind=link}